Severe Tropical Cyclone Larry
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Brooklyn, Cloudland, Melsonby (Gaarraay)
BUSH BLITZ SPECIES DISCOVERY PROGRAM Brooklyn, Cloudland, Melsonby (Gaarraay) Nature Refuges Eubenangee Swamp, Hann Tableland, Melsonby (Gaarraay) National Parks Upper Bridge Creek Queensland 29 April–27 May · 26–27 July 2010 Australian Biological Resources Study What is Contents Bush Blitz? Bush Blitz is a four-year, What is Bush Blitz? 2 multi-million dollar Abbreviations 2 partnership between the Summary 3 Australian Government, Introduction 4 BHP Billiton and Earthwatch Reserves Overview 6 Australia to document plants Methods 11 and animals in selected properties across Australia’s Results 14 National Reserve System. Discussion 17 Appendix A: Species Lists 31 Fauna 32 This innovative partnership Vertebrates 32 harnesses the expertise of many Invertebrates 50 of Australia’s top scientists from Flora 62 museums, herbaria, universities, Appendix B: Threatened Species 107 and other institutions and Fauna 108 organisations across the country. Flora 111 Appendix C: Exotic and Pest Species 113 Fauna 114 Flora 115 Glossary 119 Abbreviations ANHAT Australian Natural Heritage Assessment Tool EPBC Act Environment Protection and Biodiversity Conservation Act 1999 (Commonwealth) NCA Nature Conservation Act 1992 (Queensland) NRS National Reserve System 2 Bush Blitz survey report Summary A Bush Blitz survey was conducted in the Cape Exotic vertebrate pests were not a focus York Peninsula, Einasleigh Uplands and Wet of this Bush Blitz, however the Cane Toad Tropics bioregions of Queensland during April, (Rhinella marina) was recorded in both Cloudland May and July 2010. Results include 1,186 species Nature Refuge and Hann Tableland National added to those known across the reserves. Of Park. Only one exotic invertebrate species was these, 36 are putative species new to science, recorded, the Spiked Awlsnail (Allopeas clavulinus) including 24 species of true bug, 9 species of in Cloudland Nature Refuge. -

Known Impacts of Tropical Cyclones, East Coast, 1858 – 2008 by Mr Jeff Callaghan Retired Senior Severe Weather Forecaster, Bureau of Meteorology, Brisbane
ARCHIVE: Known Impacts of Tropical Cyclones, East Coast, 1858 – 2008 By Mr Jeff Callaghan Retired Senior Severe Weather Forecaster, Bureau of Meteorology, Brisbane The date of the cyclone refers to the day of landfall or the day of the major impact if it is not a cyclone making landfall from the Coral Sea. The first number after the date is the Southern Oscillation Index (SOI) for that month followed by the three month running mean of the SOI centred on that month. This is followed by information on the equatorial eastern Pacific sea surface temperatures where: W means a warm episode i.e. sea surface temperature (SST) was above normal; C means a cool episode and Av means average SST Date Impact January 1858 From the Sydney Morning Herald 26/2/1866: an article featuring a cruise inside the Barrier Reef describes an expedition’s stay at Green Island near Cairns. “The wind throughout our stay was principally from the south-east, but in January we had two or three hard blows from the N to NW with rain; one gale uprooted some of the trees and wrung the heads off others. The sea also rose one night very high, nearly covering the island, leaving but a small spot of about twenty feet square free of water.” Middle to late Feb A tropical cyclone (TC) brought damaging winds and seas to region between Rockhampton and 1863 Hervey Bay. Houses unroofed in several centres with many trees blown down. Ketch driven onto rocks near Rockhampton. Severe erosion along shores of Hervey Bay with 10 metres lost to sea along a 32 km stretch of the coast. -

Summary of 2005/6 Australian-Region Tropical Storm Season and Verification of Authors’ Seasonal Forecasts
Summary of 2005/6 Australian-Region Tropical Storm Season and Verification of Authors’ Seasonal Forecasts Issued: 15th May 2006 by Professor Mark Saunders and Dr Adam Lea Benfield Hazard Research Centre, UCL (University College London), UK. Summary ¢¡¤£¦¥¨§¨§¨© £¨¨ £¨£¨ ¨£"!$#%¨&('(¨)¨'*+%,¨-'(.¨&/%¨01¨.%'23'(%546¨7+£¨ ¤£¨ ¨£8!$#%¨&('(¨9&*¨¤7:¨&(&*'( ;¨.%'23'*%54=<3 ¢¡¤'*>.¨¤%¨%?@'(%¡9%¡£A:B¨#A-,£C3'(¨#/!$#%¨&('(¨ %,¨-¤'*.¨&%¨0D£¨¨E@¡'(.¡F¨&(&3¡¤¨7F)£&(,@£¨ ¨£"¨.%'23'(%54¨<G ¢¡£" ¢HIJ%£,.'(&(£K-,¨)¤¨)'(&*'(%54 :B¨,£.¨%¦:L¨M%¡¤£¥¨§¨§¨© N £¨¨O¡,@£7P£Q¨.£&*&(£%$R¤'*&(&(ST-,£7'(.%'( :,¨0VUW 4X¥¨§¨§¨©+%¡¨% !K#%¨&('*¨ )¨'( ¨7 &(¨7¤:¨&*&('( Y¨.%'Z3'(%54Y[¨#&(7 )£\.&*¨£\%Y£¨ ¨£<X ¢¡¤£] ¢H¤I 7¤£%£0^'*¤'*%'*.¦:B¨,£.¨%A¡,@£76 ¨¨¨76R¤'*&(&_:B¨;)¤¨'(`¨.%'23'(%54a¨76:B¨¦&*¨¤7:¨&(&*'( M%,¨-'(.¨& %¨0b¤#0^)£_:,¨0bUc 4d¥¨§¨§¨©¨< egf3hejilk m3n(opqsr=t k iu1vKn(wyxOze{r|vK}$ok ~3wk it nZ¨u1m¨ilhwh~¨tNw@pFwy¨u[upiOk¨st(f3h¨¨¨ ^"3wyt(ilpq(n*p~3 ilh,¨n(k ~;t*ilk m3n(opqwtNk iyuwhpwk ~9p~3¦p8gpq(n(¨pt n(k ~k¨t(f3hniwhpwk ~3pqm¨ilk 3p3n(q*n(wyt n(oAp~3¦¨ht hiunZ~3n(wyt n*o k ilhopwyt w>k i?t(f3n(w>potNn 3nZt*¨¤egf3hwh$k ilhopwyt w{@hilh$n*wwy3hKuk ~¨t(f3qZ95ilk ut*f3h$ t(f¦Wp9¨¨¨ "tNk"t*f3h t(f@hohu[3hi8¨¨¨ p~3nZ~3oqZ3¨hKwhm3pilptNhTm¨ilh¨n(otNn(k ~3wsk i{~¨¨u3hilwsk¨¤t*ilk m3n(opqwyt k iuw~¨¨u[3hilw k¨¢wh¤hilh t(ilk m3n(opq¡oC3oq(k ~3hw"p~3¢t(f3h A£E¤¥z5@oo¨u3q(ptNh^£E3oq*k ~3h¦¤¡~3hi 3}snZ~3¨h§3egf3h[q*pt(tNhiKn(w ¨n ¤h~dk i¨t*f3h/"f3k¨q(h/"3wyt(ilpq*n(p~35ilh ¨n(k ~3 Features of the 2005/6 Australian-Region Season • The 2005/6 Australian-region tropical storm season featured 11 storms of which 7 made severe tropical cyclone strength (U.S. -

In the Aftermath of Cyclone Yasi: AIR Damage Survey Observations
AIRCURRENTS By IN THE AFTERMATH OF CYCLONE YaSI: AIR DAMAGE SURVEY OBSERVATIONS EDITOR’S NOTE: In the days following Yasi’s landfall, AIR’s post-disaster survey team visited areas in Queensland affected by the storm. This article 03.2011 presents their findings. By Dr. Kyle Butler and Dr. Vineet Jain Edited by Virginia Foley INTRODUCTION Severe Cyclone Yasi began as a westward-moving depression In addition to the intense winds, Yasi brought 200-300 off Fiji that rapidly developed into a tropical storm before millimeters of precipitation in a 24 hour period and a storm dawn on January 30, 2011. Hours later, Yasi blew across the surge as high as five meters near Mission Beach. northern islands of Vanuatu, continuing to grow in intensity and size and prompting the evacuation of more than 30,000 residents in Queensland, Australia. By February 2 (local time), the storm had achieved Category 5 status on the Australian cyclone scale (a strong Category 4 on the Saffir-Simpson scale). At its greatest extent, the storm spanned 650 kilometers. By the next day, it became clear that Yasi would spare central Queensland, which had been devastated by heavy flooding in December. But there was little else in the way of good news. Yasi made landfall on the northeast coast of Queensland on February 3 between Innisfail and Cardwell with recorded gusts of 185 km/h. Satellite-derived sustained Figure 1. Satellite-derived 10-minute sustained wind speeds in knots just after wind speeds of 200 km/h were estimated near the center Cyclone Yasi made landfall. -

Take Another Look
Take Contact Details Another SUNSHINE COAST REGIONAL COUNCIL Caloundra Customer Service Look..... 1 Omrah Avenue, Caloundra FRONT p: 07 5420 8200 e: [email protected] Maroochydore Customer Service 11-13 Ocean Street, Maroochydore p: 07 5475 8501 e: [email protected] Nambour Customer Service Cnr Currie & Bury Street, Nambour p: 07 5475 8501 e: [email protected] Tewantin Customer Service 9 Pelican Street, Tewantin p: 07 5449 5200 e: [email protected] YOUR LOCAL CONTACT Our Locals are Beauties HINTERLAND EDITION HINTERLAND EDITION 0 Local native plant guide 2 What you grow in your garden can have major impact, Introduction 3 for better or worse, on the biodiversity of the Sunshine Native plants 4 - 41 Coast. Growing a variety of native plants on your property can help to attract a wide range of beautiful Wildlife Gardening 20 - 21 native birds and animals. Native plants provide food and Introduction Conservation Partnerships 31 shelter for wildlife, help to conserve local species and Table of Contents Table Environmental weeds 42 - 73 enable birds and animals to move through the landscape. Method of removal 43 Choosing species which flower and fruit in different Succulent plants and cacti 62 seasons, produce different types of fruit and provide Water weeds 70 - 71 roost or shelter sites for birds, frogs and lizards can greatly increase your garden’s real estate value for native References and further reading 74 fauna. You and your family will benefit from the natural pest control, life and colour that these residents and PLANT TYPE ENVIRONMENTAL BENEFITS visitors provide – free of charge! Habitat for native frogs Tall Palm/Treefern Local native plants also improve our quality of life in Attracts native insects other ways. -

MARCH 2021 SGAP Revisits Babinda Golf Course
NEWSLETTER 208 MARCH 2021 SGAP revisits Babinda Golf Course Don Lawie Our first excursion for the new year was a return to the green field of Babinda Golf Club. The height of the wet season was upon us and we looked for a site that was botanically interesting and had shelter in case of rain. Babinda, Australia’s wettest town, is well set up for rainy days and we were welcomed by Golf Club members Peter and Patsy who are also SGAP members. On our visit in [Editors note: uncountable years ago] we had to to dodge the golfers as they played a round but today the god of rain had performed an apotropaic release flowing drains. The fairways are timber tree and suffering from an from their sysiphean task and we had delineated by rows of single trees, attack of myrtle rust. A notable the course to ourselves. About fifteen almost all of which are species native specimen, not native to the Babinda of us enjoyed a leisurely lunch and a to the area, supplied by native plant area, was possibly Austromullera valida, discussion of plants on the specimen enthusiasts including Nigel Tucker and from the high country of Mt Lewis, table. Stuart displayed a magnificent Rob Jago. They were planted about home of many rarities. metre long stem of Banksia robur with thirty years ago and are a lesson in two large inflorescences, a small piece how rainforest trees will grow when of fruit- bearing Finger Lime and a not associated with the close growth flowering Brachychiton vitifolius stem of their natural habitat. -

Lessons Learned from Cyclones in Northern Australia
Lessons Learned from Cyclones in Northern Australia SEPTEMBER 2013 RIRDC Publication No. 13/071 Lessons Learned from Cyclones in Northern Australia By Rhonda Sorensen September 2013 RIRDC Publication No. 13/071 RIRDC Project No. PRJ-008498 © 2013 Rural Industries Research and Development Corporation. All rights reserved. ISBN 978-1-74254-564-6 ISSN 1440-6845 Lessons Learned from Cyclones in Northern Australia Publication No. 13/071 Project No. PRJ-008498 The information contained in this publication is intended for general use to assist public knowledge and discussion and to help improve the development of sustainable regions. You must not rely on any information contained in this publication without taking specialist advice relevant to your particular circumstances. While reasonable care has been taken in preparing this publication to ensure that information is true and correct, the Commonwealth of Australia gives no assurance as to the accuracy of any information in this publication. The Commonwealth of Australia, the Rural Industries Research and Development Corporation (RIRDC), the authors or contributors expressly disclaim, to the maximum extent permitted by law, all responsibility and liability to any person, arising directly or indirectly from any act or omission, or for any consequences of any such act or omission, made in reliance on the contents of this publication, whether or not caused by any negligence on the part of the Commonwealth of Australia, RIRDC, the authors or contributors. The Commonwealth of Australia does not necessarily endorse the views in this publication. This publication is copyright. Apart from any use as permitted under the Copyright Act 1968, all other rights are reserved. -

The Cyclone As Trope of Apocalypse and Place in Queensland Literature
ResearchOnline@JCU This file is part of the following work: Spicer, Chrystopher J. (2018) The cyclone written into our place: the cyclone as trope of apocalypse and place in Queensland literature. PhD Thesis, James Cook University. Access to this file is available from: https://doi.org/10.25903/7pjw%2D9y76 Copyright © 2018 Chrystopher J. Spicer. The author has certified to JCU that they have made a reasonable effort to gain permission and acknowledge the owners of any third party copyright material included in this document. If you believe that this is not the case, please email [email protected] The Cyclone Written Into Our Place The cyclone as trope of apocalypse and place in Queensland literature Thesis submitted by Chrystopher J Spicer M.A. July, 2018 For the degree of Doctor of Philosophy College of Arts, Society and Education James Cook University ii Acknowledgements of the Contribution of Others I would like to thank a number of people for their help and encouragement during this research project. Firstly, I would like to thank my wife Marcella whose constant belief that I could accomplish this project, while she was learning to live with her own personal trauma at the same time, encouraged me to persevere with this thesis project when the tide of my own faith would ebb. I could not have come this far without her faith in me and her determination to journey with me on this path. I would also like to thank my supervisors, Professors Stephen Torre and Richard Landsdown, for their valuable support, constructive criticism and suggestions during the course of our work together. -

IFE and Wlldll ITAT of AMERICAN SA VIRONMENT and ECOLO
IFE AND WlLDLl ITAT OF AMERICAN SA VIRONMENT AND ECOLO By A. Binion Amerson, Jr., W. Arthur Whistler, and Terry D. Schwaner Environment Consultants, Inc., Dallas, Texas Edited by Richard C. Banks U.S. Fish and Wildlife Service Washington, D.C. UNITED STATES DEPARTMENT OF T E INTERIOR FISH AND WILDLIFE SERVICE Washington, D.C. e 1982 Foreword A survey of the status of the wildlife and wildlife habitat of American Samoa. an unincorporated Territory of the United States. was recommended by administrative officials of the U.S. Fish and Wildlife Service (FWS) in the early 1970s .Environ- ment Consultants. Inc . (ECI). based in Dallas. Texas. was selected to conduct a 2-year survey with A .Binion Amerson. Jr., as Principal Investigator . The contract was administered through the Division of Federal Aid in FWS Region I. Portland. Oregon . The primary objectives of the survey were (1) to define the major ecosystems and to inventory their physical components. vegetation. and wildlife constituents; (2) to prepare maps of these ecosystems; (3) to identify any threatened or endangered species of wildlife; and (4) to recommend wildlife management opportunities and needs . The report of the survey was to be in two parts . The first was to be a non-technical account suitable for wide general distribution; the second was to include the technical aspects of the data and data gathering. with accounts of the wildlife species . This volume represents the first part of ECI's report . The final report submitted by ECI contained more than 1. 200 pages. 200 figures. and 110 tables. many of thelatter several pages long . -

Weed Response to Cyclones in the Wet Tropics Rainforests – Impacts and Adaptation –
Weed Response to Cyclones in the Wet Tropics Rainforests – Impacts and adaptation – Pub. No. 11/010 www.rirdc.gov.au Weed response to cyclones in the Wet Tropics rainforests Impacts and adaptation by Helen T Murphy, Dan J Metcalfe, Matt G Bradford and Andrew J Ford March 2011 RIRDC Publication No 11/010 RIRDC Project No AWRC 08-61 © 2011 Rural Industries Research and Development Corporation. All rights reserved. ISBN 978-1-74254-190-0 ISSN 1440-6845 Weed response to cyclones in the Wet Tropics rainforests: impacts and adaptation Publication No. 11/010 Project No. AWRC 08-61 The information contained in this publication is intended for general use to assist public knowledge and discussion and to help improve the development of sustainable regions. You must not rely on any information contained in this publication without taking specialist advice relevant to your particular circumstances. While reasonable care has been taken in preparing this publication to ensure that information is true and correct, the Commonwealth of Australia gives no assurance as to the accuracy of any information in this publication. The Commonwealth of Australia, the Rural Industries Research and Development Corporation and the authors or contributors expressly disclaim, to the maximum extent permitted by law, all responsibility and liability to any person arising directly or indirectly from any act or omission or for any consequences of any such act or omission made in reliance on the contents of this publication, whether or not caused by any negligence on the part of the Commonwealth of Australia, RIRDC, the authors or contributors. -

Endiandric Acid Derivatives and Other Constituents of Plants from the Genera Beilschmiedia and Endiandra (Lauraceae)
Biomolecules 2015, 5, 910-942; doi:10.3390/biom5020910 OPEN ACCESS biomolecules ISSN 2218-273X www.mdpi.com/journal/biomolecules/ Review Endiandric Acid Derivatives and Other Constituents of Plants from the Genera Beilschmiedia and Endiandra (Lauraceae) Bruno Ndjakou Lenta 1,2,*, Jean Rodolphe Chouna 3, Pepin Alango Nkeng-Efouet 3 and Norbert Sewald 2 1 Department of Chemistry, Higher Teacher Training College, University of Yaoundé 1, P.O. Box 47, Yaoundé, Cameroon 2 Organic and Bioorganic Chemistry, Chemistry Department, Bielefeld University, P.O. Box 100131, 33501 Bielefeld, Germany; E-Mail: [email protected] 3 Department of Chemistry, University of Dschang, P.O. Box 67, Dschang, Cameroon; E-Mails:[email protected] (J.R.C.); [email protected] (P.A.N.-E.) * Author to whom correspondence should be addressed; E-Mail: [email protected]; Tel.: +2376-7509-7561. Academic Editor: Jürg Bähler Received: 3 March 2015 / Accepted: 6 May 2015 / Published: 14 May 2015 Abstract: Plants of the Lauraceae family are widely used in traditional medicine and are sources of various classes of secondary metabolites. Two genera of this family, Beilschmiedia and Endiandra, have been the subject of numerous investigations over the past decades because of their application in traditional medicine. They are the only source of bioactive endiandric acid derivatives. Noteworthy is that their biosynthesis contains two consecutive non-enzymatic electrocyclic reactions. Several interesting biological activities for this specific class of secondary metabolites and other constituents of the two genera have been reported, including antimicrobial, enzymes inhibitory and cytotoxic properties. This review compiles information on the structures of the compounds described between January 1960 and March 2015, their biological activities and information on endiandric acid biosynthesis, with 104 references being cited. -
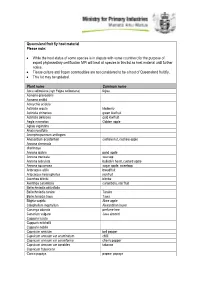
MPI Queensland Fruit Fly Host Material List
Queensland fruit fly host material Please note: While the host status of some species is in dispute with some countries; for the purpose of export phytosanitary certification MPI will treat all species in this list as host material until further notice. Tissue culture and frozen commodities are not considered to be a host of Queensland fruit fly. This list may be updated. Plant name Common name Acca sellowiana (syn Feijoa sellowiana) feijoa Acmena graveolens Acmena smithii Acroychia acidula Actinidia arguta kiwiberry Actinidia chinensis green kiwifruit Actinidia deliciosa gold kiwifruit Aegle marmelos Golden apple Aglaia sapindina Alyxia ruscifolia Amorphospermum antilogum Anacardium occidentale cashew nut, cashew apple Annona cherimola cherimoya Annona glabra pond apple Annona muricata soursop Annona reticulata bullock's heart, custard apple Annona squamosa sugar apple, sweetsop Artocarpus altilis breadfruit Artocarpus heterophyllus jackfruit Averrhoa bilimbi blimbe Averrhoa carambola carambola, star fruit Beilschmiedia obtusifolia Beilschmiedia taraire Taraire Beilschmiedia tawa Tawa Blighia sapida Akee apple Calophyllum inophyllum Alexandrian laurel Cananga odorata perfume tree Canarium vulgare Java almond Capparis lucida Capparis mitchellii Capparis nobilis Capsicum annuum bell pepper Capsicum annuum var acuminatum chilli Capsicum annuum var cerasiforme cherry pepper Capsicum annuum var conoides tabasco Capsicum frutescens Carica papaya papaw, papaya Carica pentagona babaco Carissa ovata Casimiroa edulis white sapote Cassine australia Castanospora alphandii Chrysophyllum cainito caimito, star apple Cissus sp Citrullus lanatus Watermelon Citrus spp. Citrus aurantiifolia West indian lime Citrus aurantium Seville orange Citrus grandis (syn maxima) pummelo Citrus hystrix kaffir lime Citrus jambhiri rough lemon Citrus latifolia Tahatian lime Citrus limetta sweet lemon tree Citrus limon lemon Citrus maxima (syn grandis) pomelo, shaddock Citrus medica citron Citrus meyeri Meyer lemon Citrus reticulata mandarin Citrus reticulata var.