Gastropoda, Pulmonata, Orculidae/Pupillidae) from The
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

(Pulmonata: Vertiginidae) and Strobilops
Records of the Hawaii Biological Survey for 2012. Edited by Neal L. Evenhuis & Lucius G. Eldredge. Bishop Museum Occasional Papers 114: 39 –42 (2013) Hawaiian land snail records : Lyropupa cookei Clench , 1952 (Pulmonata : Vertiginidae ) and Strobilops aeneus Pilsbry , 1926 (Pulmonata : Strobilopsidae ) CARl C. C HRiSTeNSeN Bishop Museum, 1525 Bernice Street, Honolulu, Hawai‘i 96817-2704, USA; email: [email protected] This note clarifies the status of two taxa of land snails that have been reported to occur in the Hawaiian islands. Lyropupa cookei Clench, 1952, is shown to be a synonym of Lyropupa anceyana Cooke & Pilsbry in Pilsbry & Cooke, 1920. The sole Hawaiian record for the North American Strobilops aeneus Pilsbry, 1926, is almost certainly based on a mislabeled specimen, and accordingly this species should be removed from the Hawaiian faunal list. Lyropupa cookei Clench , 1952 Lyropupa Pilsbry, 1900, is a genus of pupilloid land snails endemic to the Hawaiian islands. in their monograph of the genus, Pilsbry & Cooke (1920 in 1918–1920: 253–254, pl. 26, figs. 3, 6) published a description of “ Lyropupa anceyana C. & P., n. sp.,” based on specimens from ola‘a on the island of Hawai‘i held in the collections of Bishop Museum and the Academy of Natural Sciences of Philadelphia. They stated that their new species had previously been misidentified by Ancey (1904:124) as Lyropupa lyrata (Gould, 1843) . Several pages earlier, in their systematic treatment of that species, Pilsbry & Cooke (1918–1920: 235) had also set forth their conclusion that Ancey had misidenti - fied Gould’s species and stated that in fact Ancey’s “description of lyrata was based on specimens of an unnamed species for which the name L. -

Phylogeography of the East-Alpine Members of the Landsnail Species Orcula Dolium (Gastropoda: Pulmonata: Orculidae)
©Institut f. Erdwissensch., Geol. u. Paläont., Karl-Franzens-Universität Graz; download www.biologiezentrum.at Phylogeography of the East-Alpine members of the landsnail species Orcula dolium (Gastropoda: Pulmonata: Orculidae) Harl, J., Kruckenhauser, L., Sattmann, H., Duda, M. & Haring, E. Museum of Natural History Vienna, Burgring 7, A-1010 Vienna, Austria, e-mail: [email protected], [email protected], [email protected], [email protected], [email protected] Orcula dolium has the widest distribution among the species of the genus Orcula and inhabits the calcareous parts of the Alps and the Northern Carpathians. Since it had been described in 1801 by DRAPARNAUD, various authors added a minimum of 23 further subspecies. Most of these subspecies represent local forms that inhabit only small, partly isolated regions and differ from the nominate subspecies in their shell shape and the expression of their aperture's folds. Besides the common O. dolium dolium, six subspecies have been described from the Eastern Alps. Two of them, O. dolium edita and O. dolium raxae, are restricted to different elevations of high mountain areas. According to the literature, the nominate subspecies is sporadically found in the same habitats as the high alpine forms, but no intermediate morphs have been detected. This observation raises the question if these taxa represent distinct species. To determine whether these taxa are differentiated genetically and to reveal their relationships, snails were collected from sites covering the main part of the East Alpine distribution range of the species. Two mitochondrial genes (COI and 16S rRNA) as well as the nuclear histone H3-H4-spacer region were sequenced and genetic distances calculated. -

Arianta 6, 2018
ZOBODAT - www.zobodat.at Zoologisch-Botanische Datenbank/Zoological-Botanical Database Digitale Literatur/Digital Literature Zeitschrift/Journal: Arianta Jahr/Year: 2018 Band/Volume: 6 Autor(en)/Author(s): diverse Artikel/Article: Abstracts Talks Alpine and other land snails 11-27 ARIANTA 6 and correspond ecologically. For instance, the common redstart is a bird species breeding in the lowlands, whereas the black redstart is native to higher altitudes. Some species such as common swift and kestrel, which are originally adapted to enduring in rocky areas, even found a secondary habitat in the house facades and street canyons of towns and big cities. Classic rock dwellers include peregrine, eagle owl, rockthrush, snowfinch and alpine swift. The presentation focuses on the biology, causes of threat as well as conservation measures taken by the national park concerning the species golden eagle, wallcreeper, crag martin and ptarmigan. Birds breeding in the rocks might not be that high in number, but their survival is all the more fascinating and worth protecting as such! Abstracts Talks Alpine and other land snails Arranged in chronological order of the program Rangeconstrained cooccurrence simulation reveals little niche partitioning among rockdwelling Montenegrina land snails (Gastropoda: Clausiliidae) Zoltán Fehér1,2,3, Katharina JakschMason1,2,4, Miklós Szekeres5, Elisabeth Haring1,4, Sonja Bamberger1, Barna PállGergely6, Péter Sólymos7 1 Central Research Laboratories, Natural History Museum Vienna, Austria; [email protected] -

(Mollusca, Gastropoda) of the Bulgarian Part of the Alibotush Mts
Malacologica Bohemoslovaca (2008), 7: 17–20 ISSN 1336-6939 Terrestrial gastropods (Mollusca, Gastropoda) of the Bulgarian part of the Alibotush Mts. IVAILO KANEV DEDOV Central Laboratory of General Ecology, 2 Gagarin Str., BG-1113 Sofia, Bulgaria, e-mail: [email protected] DEDOV I.K., 2008: Terrestrial gastropods (Mollusca, Gastropoda) of the Bulgarian part of the Alibotush Mts. – Malacologica Bohemoslovaca, 7: 17–20. Online serial at <http://mollusca.sav.sk> 20-Feb-2008. This work presents results of two years collecting efforts within the project “The role of the alpine karst area in Bulgaria as reservoir of species diversity”. It summarizes distribution data of 44 terrestrial gastropods from the Bulgarian part of Alibotush Mts. Twenty-seven species are newly recorded from the Alibotush Mts., 13 were con- firmed, while 4 species, previously known from the literature, were not found. In the gastropod fauna of Alibotush Mts. predominate species from Mediterranean zoogeographic complex. A large part of them is endemic species, and this demonstrates the high conservation value of large limestone areas in respect of terrestrial gastropods. Key words: terrestrial gastropods, distribution, Alibotush Mts., Bulgaria Introduction Locality 6: vill. Katuntsi, Izvorite hut, near hut, open The Alibotush Mts. (other popular names: Kitka, Gotseva ruderal terrain, under bark, 731 m a.s.l., coll. I. Dedov. Planina, Slavjanka) is one of the most interesting large Locality 7: vill. Katuntsi, tufa-gorge near village, 700 m limestone area in Bulgaria (Fig. 1). It occupies the part a.s.l., coll. I. Dedov, N. Simov. of the border region between Bulgaria and Greece with Locality 8: below Livade area, road between Goleshevo maximum elevation 2212 m (Gotsev peak). -

Pupillid Land Snails of Eastern North America*
Amer. Malac. Bull. 28: 1-29 (2010) Pupillid land snails of eastern North America* Jeffrey C. Nekola1 and Brian F. Coles2 1 Department of Biology, University of New Mexico, Albuquerque, New Mexico 87131, U.S.A. 2 Mollusca Section, Department of Biodiversity, National Museum of Wales, Cathays Park, Cardiff CF10 3NP, U.K. Corresponding author: [email protected] Abstract: The Pupillidae form an important component of eastern North American land snail biodiversity, representing approx. 12% of the entire fauna, 25-75% of all species and individuals at regional scales, at least 30% of the species diversity, and 33% of individuals within any given site. In some regions pupillids represent 80-100% of total molluscan diversity within sites, notably in taiga, tundra, and the base-poor pine savannas and pocosins of the southeastern coastal plain. Adequate documentation of North American land snail biodiversity thus requires investigators to effi ciently collect and accurately identify individuals of this group. This paper presents a set of annotated keys to the 65 species in this family known to occur in North America east of the Rocky Mountains. The distinguishing taxonomic features, updated county-scale range maps, and ecological conditions favored by each are presented in hopes of stimulating future research in this important group. Key words: microsnail, biodiversity, ecology, biogeography, taxonomy For the last dozen years, our interests in terrestrial Adequate documentation of this diversity thus requires gastropod biodiversity have lead us individually and investigators to effi ciently collect and accurately identify collectively to observe molluscan communities over most of individuals from this family. Unfortunately, neither has been North America, ranging from central Quebec, Hudson’s Bay common. -

Liste Rouge Mollusques (Gastéropodes Et Bivalves)
2012 > L’environnement pratique > Listes rouges / Gestion des espèces > Liste rouge Mollusques (gastéropodes et bivalves) Espèces menacées en Suisse, état 2010 > L’environnement pratique > Listes rouges / Gestion des espèces > Liste rouge Mollusques (gastéropodes et bivalves) Espèces menacées en Suisse, état 2010 Publié par l’Office fédéral de l’environnement OFEV et par le Centre suisse de cartographie de la faune CSCF Berne, 2012 Valeur juridique de cette publication Impressum Liste rouge de l’OFEV au sens de l’art. 14, al. 3, de l’ordonnance Editeurs du 16 janvier 1991 sur la protection de la nature et du paysage Office fédéral de l’environnement (OFEV) (OPN; RS 451.1), www.admin.ch/ch/f/rs/45.html L’OFEV est un office du Département fédéral de l’environnement, des transports, de l’énergie et de la communication (DETEC). La présente publication est une aide à l’exécution de l’OFEV en tant Centre Suisse de Cartographie de la Faune (CSCF), Neuchâtel. qu’autorité de surveillance. Destinée en premier lieu aux autorités d’exécution, elle concrétise des notions juridiques indéterminées Auteurs provenant de lois et d’ordonnances et favorise ainsi une application Mollusques terrestres: Jörg Rüetschi, Peter Müller et François Claude uniforme de la législation. Elle aide les autorités d’exécution Mollusques aquatiques: Pascal Stucki et Heinrich Vicentini notamment à évaluer si un biotope doit être considéré comme digne avec la collaboration de Simon Capt et Yves Gonseth (CSCF) de protection (art. 14, al. 3, let. d, OPN). Accompagnement à l’OFEV Francis Cordillot, division Espèces, écosystèmes, paysages Référence bibliographique Rüetschi J., Stucki P., Müller P., Vicentini H., Claude F. -

Spatial Predictive Distribution Modelling of Madeira's Endemic
DEPARTAMENTO DE ZOOLOGIA FACULDADE DE CIÊNCIAS E TECNOLOGIA UNIVERSIDADE DE COIMBRA Spatial predictive distribution modelling of Madeira’s endemic land snail species Dinarte Nuno Freitas Teixeira 2009 REGIÃO AUTÓNOMA DA REPÚBLICA PORTUGUESA UNIÃO EUROPEIA MADEIRA FSE DEPARTAMENTO DE ZOOLOGIA FACULDADE DE CIÊNCIAS E TECNOLOGIA UNIVERSIDADE DE COIMBRA Spatial predictive distribution modelling of Madeira’s endemic land snail species Dissertação apresentada à Universidade de Coimbra para cumprimento dos requisitos necessários à obtenção do grau de Mestre em Ecologia, realizada sob a orientação científica do Professor Doutor José Paulo Sousa (Universidade de Coimbra) e do Professor Doutor José Manuel Jesus (Universidade da Madeira). Dinarte Nuno Freitas Teixeir a 2009 O presente trabalho foi financiado pelo Centro de Ciência e Tecnologia da Madeira (CITMA), através da bolsa de Mestrado FSE BM I/2008 – 531, ao abrigo do Programa Operacional de Valorização do Potencial Humano e Coesão Social da RAM (RUMOS). REGIÃO AUTÓNOMA DA MADEIRA REPÚBLICA PORTUGUESA UNIÃO EUROPEIA FSE À Susana AGRADECIMENTOS Esta tese é o resultado de um trabalho conjunto para o qual muitos contribuíram e aos quais desejo reconhecer e deixar o meu agradecimento. Ao professor Doutor José Paulo Sousa, meu orientador, pela indispensável ajuda, paciência e orientação científica. Ao professor Doutor José Manuel Jesus, meu orientador, pela amizade e apoio desde os primeiros momentos. Pelo seu empenho, conselhos transmitidos, chamadas à razão e orientação científica o meu muito obrigado. Ao Doutor Pedro Cardoso, meu orientador e a quem muito devo, pelo constante acompanhamento e disponibilidade, amizade e orientação científica. Por tudo o que me ensinou, pela motivação e animo que sempre me transmitiu, e, acima de tudo, pela manutenção da objectividade do trabalho. -
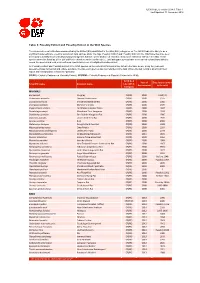
Table 9: Possibly Extinct and Possibly Extinct in the Wild Species
IUCN Red List version 2014.3: Table 9 Last Updated: 13 November 2014 Table 9: Possibly Extinct and Possibly Extinct in the Wild Species The number of recent extinctions documented by the Extinct (EX) and Extinct in the Wild (EW) categories on The IUCN Red List is likely to be a significant underestimate, even for well-known taxa such as birds. The tags 'Possibly Extinct' and 'Possibly Extinct in the Wild' have therefore been developed to identify those Critically Endangered species that are, on the balance of evidence, likely to be extinct (or extinct in the wild). These species cannot be listed as EX or EW until their extinction can be confirmed (i.e., until adequate surveys have been carried out and have failed to record the species and local or unconfirmed reports have been investigated and discounted). All 'Possibly Extinct' and 'Possibly Extinct in the Wild' species on the current IUCN Red List are listed in the table below, along the year each assessment was carried out and, where available, the date each species was last recorded in the wild. Where the last record is an unconfirmed report, last recorded date is noted as "possibly". CR(PE) - Critically Endangered (Possibly Extinct), CR(PEW) - Critically Endangered (Possibly Extinct in the Wild), IUCN Red Year of Date last recorded Scientific name Common name List (2014) Assessment in the wild Category MAMMALS Bos sauveli Kouprey CR(PE) 2008 1969/70 Crateromys australis Dinagat Crateromys CR(PE) 2008 1975 Crocidura trichura Christmas Island Shrew CR(PE) 2008 1985 Crocidura wimmeri -

Strobilops Aeneus Pilsbry, 1926, in Canada, with Two New Ontario Records (Mollusca: Gastropoda
Check List 10(2): 397–401, 2014 © 2014 Check List and Authors Chec List ISSN 1809-127X (available at www.checklist.org.br) Journal of species lists and distribution N Distribution of Strobilops aeneus Pilsbry, 1926, in Canada, with two new Ontario records (Mollusca: Gastropoda: ISTRIBUTIO Strobilopsidae) D 1* 2 RAPHIC Robert G. Forsyth , and Michael J. Oldham G EO G 1 New Brunswick Museum, 277 Douglas Avenue, Saint John, New Brunswick, Canada E2K 1E5 N 2 Natural Heritage Information Centre, Ministry of Natural Resources, 300 Water Street, PO Box 7000, Peterborough, Ontario, Canada K9J 8M5 O * Corresponding author. E-mail: [email protected] OTES N Abstract: The geographic distribution of Strobilops aeneus Pilsbry, 1926, a rare species in Canada, is reviewed and all known records are mapped. Two recent records, the only ones since 1941, are reported from the province of Ontario. One of these records represents a small range extension ca S. aeneus from Ontario, New Brunswick, and Nova Scotia were re-examined and found to be another species. 85 km north of the closest previous site. Specimens identified as There are just over 200 species of land snails and from Cape Breton Island, Nova Scotia, and by Lauriol slugs in Canada, and while the majority of species are et al. (2003) in a palaeoecological study of Holocene broad Nearctic or Holarctic ranges, others are range-edge species that occur in Canada just north of the border with southwestern Quebec. There are no published records of the United States (Forsyth 2013). Southern Ontario has thecave species infill on from the New Eardley Brunswick, Escarpment, but the Gatineau New Brunswick Park, in special malacological interest because of a concentration of range-edge species that in Canada only occur here and have restricted ranges (Pilsbry 1940, 1946, 1948; Oughton andMuseum a key had to Strobilopsone record species. -

CLECOM-Liste Österreich (Austria)
CLECOM-Liste Österreich (Austria), mit Änderungen CLECOM-Liste Österreich (Austria) Phylum Mollusca C UVIER 1795 Classis Gastropoda C UVIER 1795 Subclassis Orthogastropoda P ONDER & L INDBERG 1995 Superordo Neritaemorphi K OKEN 1896 Ordo Neritopsina C OX & K NIGHT 1960 Superfamilia Neritoidea L AMARCK 1809 Familia Neritidae L AMARCK 1809 Subfamilia Neritinae L AMARCK 1809 Genus Theodoxus M ONTFORT 1810 Subgenus Theodoxus M ONTFORT 1810 Theodoxus ( Theodoxus ) fluviatilis fluviatilis (L INNAEUS 1758) Theodoxus ( Theodoxus ) transversalis (C. P FEIFFER 1828) Theodoxus ( Theodoxus ) danubialis danubialis (C. P FEIFFER 1828) Theodoxus ( Theodoxus ) danubialis stragulatus (C. P FEIFFER 1828) Theodoxus ( Theodoxus ) prevostianus (C. P FEIFFER 1828) Superordo Caenogastropoda C OX 1960 Ordo Architaenioglossa H ALLER 1890 Superfamilia Cyclophoroidea J. E. G RAY 1847 Familia Cochlostomatidae K OBELT 1902 Genus Cochlostoma J AN 1830 Subgenus Cochlostoma J AN 1830 Cochlostoma ( Cochlostoma ) septemspirale septemspirale (R AZOUMOWSKY 1789) Cochlostoma ( Cochlostoma ) septemspirale heydenianum (C LESSIN 1879) Cochlostoma ( Cochlostoma ) henricae henricae (S TROBEL 1851) - 1 / 36 - CLECOM-Liste Österreich (Austria), mit Änderungen Cochlostoma ( Cochlostoma ) henricae huettneri (A. J. W AGNER 1897) Subgenus Turritus W ESTERLUND 1883 Cochlostoma ( Turritus ) tergestinum (W ESTERLUND 1878) Cochlostoma ( Turritus ) waldemari (A. J. W AGNER 1897) Cochlostoma ( Turritus ) nanum (W ESTERLUND 1879) Cochlostoma ( Turritus ) anomphale B OECKEL 1939 Cochlostoma ( Turritus ) gracile stussineri (A. J. W AGNER 1897) Familia Aciculidae J. E. G RAY 1850 Genus Acicula W. H ARTMANN 1821 Acicula lineata lineata (DRAPARNAUD 1801) Acicula lineolata banki B OETERS , E. G ITTENBERGER & S UBAI 1993 Genus Platyla M OQUIN -TANDON 1856 Platyla polita polita (W. H ARTMANN 1840) Platyla gracilis (C LESSIN 1877) Genus Renea G. -
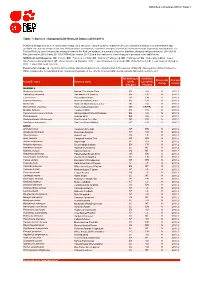
Table 7: Species Changing IUCN Red List Status (2010-2011)
IUCN Red List version 2011.2: Table 7 Table 7: Species changing IUCN Red List Status (2010-2011) Published listings of a species' status may change for a variety of reasons (genuine improvement or deterioration in status; new information being available that was not known at the time of the previous assessment; taxonomic changes; corrections to mistakes made in previous assessments, etc. To help Red List users interpret the changes between the Red List updates, a summary of species that have changed category between 2010 (IUCN Red List version 2010.4) and 2011 (IUCN Red List version 2011.2) and the reasons for these changes is provided in the table below. IUCN Red List Categories: EX - Extinct, EW - Extinct in the Wild, CR - Critically Endangered, EN - Endangered, VU - Vulnerable, LR/cd - Lower Risk/conservation dependent, NT - Near Threatened (includes LR/nt - Lower Risk/near threatened), DD - Data Deficient, LC - Least Concern (includes LR/lc - Lower Risk, least concern). Reasons for change: G - Genuine status change (genuine improvement or deterioration in the species' status); N - Non-genuine status change (i.e., status changes due to new information, improved knowledge of the criteria, incorrect data used previously, taxonomic revision, etc.) IUCN Red List IUCN Red Reason for Red List Scientific name Common name (2010) List (2011) change version Category Category MAMMALS Bradypus torquatus Maned Three-toed Sloth EN VU N 2011.1 Callicebus oenanthe San Martin Titi Monkey EN CR N 2011.1 Equus ferus Przewalski's Horse CR EN G 2011.2 -

European Glacial Relict Snails and Plants: Environmental Context of Their Modern Refugial Occurrence in Southern Siberia
bs_bs_banner European glacial relict snails and plants: environmental context of their modern refugial occurrence in southern Siberia MICHAL HORSAK, MILAN CHYTRY, PETRA HAJKOV A, MICHAL HAJEK, JIRI DANIHELKA, VERONIKA HORSAKOV A, NIKOLAI ERMAKOV, DMITRY A. GERMAN, MARTIN KOCI, PAVEL LUSTYK, JEFFREY C. NEKOLA, ZDENKA PREISLEROVA AND MILAN VALACHOVIC Horsak, M., Chytry, M., Hajkov a, P., Hajek, M., Danihelka, J., Horsakov a,V.,Ermakov,N.,German,D.A.,Ko cı, M., Lustyk, P., Nekola, J. C., Preislerova, Z. & Valachovic, M. 2015 (October): European glacial relict snails and plants: environmental context of their modern refugial occurrence in southern Siberia. Boreas, Vol. 44, pp. 638–657. 10.1111/bor.12133. ISSN 0300-9483. Knowledge of present-day communities and ecosystems resembling those reconstructed from the fossil record can help improve our understanding of historical distribution patterns and species composition of past communities. Here, we use a unique data set of 570 plots explored for vascular plant and 315 for land-snail assemblages located along a 650-km-long transect running across a steep climatic gradient in the Russian Altai Mountains and their foothills in southern Siberia. We analysed climatic and habitat requirements of modern populations for eight land-snail and 16 vascular plant species that are considered characteristic of the full-glacial environment of central Europe based on (i) fossil evidence from loess deposits (snails) or (ii) refugial patterns of their modern distribu- tions (plants). The analysis yielded consistent predictions of the full-glacial central European climate derived from both snail and plant populations. We found that the distribution of these 24 species was limited to the areas with mean annual temperature varying from À6.7 to 3.4 °C (median À2.5 °C) and with total annual precipitation vary- ing from 137 to 593 mm (median 283 mm).