Antiatherogenic Effect of Probucol Unrelated to Its Can Selectively Inhibit Low Density Lipoprotein Degradation in Atheroscleros
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Deliverable 5.A Interim Report on the Study Results APPENDIX 2
Deliverable 5.a Interim report on the study results APPENDIX 2: Algorithms used to identify study variables for service contract EMA/2011/38/CN ‐ PIOGLITAZONE November 28th 2012 D5.a Interim report on the study results (Appendix 2) for Service Contract EMA/2011/38/CN PIOGLITAZONE Author(s): Vera Ehrenstein (AUH‐AS) APPENDIX 2. ALGORITHMS USED TO IDENTIFY STUDY VARIABLES Algorithms for AU Database DISEASE/CONDITION ICD-8 CODE (1977-1993) ICD-10 CODE (1994-) Diabetes type 2 250.00; 250.06; 250.07; 250.09 E11.0; E11.1; E11.9 Cancer of bladder 188 C67 Haematuria N/A R31 Haematuria, unspecified B18, K70.0–K70.3, K70.9, K71, K73, Mild hepatic impairment 571, 573.01, 573.04 K74, K76.0 Moderate to severe hepatic 070.00, 070.02, 070.04, 070.06, B15.0, B16.0, B16.2, B19.0, K70.4, impairment 070.08, 573.00, 456.00–456.09 K72, K76.6, I85 Acute myocardial infarction 410 I21-I23 Acute coronary syndrome 410, 413 I20-I24 Ischemic heart disease 410-414 I20-I25 427.09, 427.10, 427.11, 427.19, Congestive heart failure I50, I11.0, I13.0,I13.2 428.99, 782.49; Acute renal failure N/A N17 Diabetic coma N/A E10.0, E11.0, E12.0,E13.0, E14.0 Diabetic acidosis N/A E10.1, E11.1, E12.1,E13.1, E14.1 F10.1-F10.9, G31.2, G62.1, G72.1, Alcoholism 291, 303, 577.10, 571.09, 571.10 I42.6, K29.2, K86.0, Z72.1 Obesity 277.99 E65-E66 D5.a Interim report on the study results (Appendix 2) for Service Contract EMA/2011/38/CN PIOGLITAZONE Author(s): Vera Ehrenstein (AUH‐AS) Algorithms for defining acute events in Denmark, ICD-10 code Event ICD-10 code I21.x, I23.x http://apps.who.int/classifications/icd10/browse/2010/en#/I21 -

Consumer Medicine Information
NEW ZEALAND DATA SHEET PRAVASTATIN MYLAN 1. Product Name Pravastatin Mylan, 10 mg, 20 mg and 40 mg, tablets 2. Qualitative and Quantitative Composition Each tablet contains either 10 mg, 20 mg or 40 mg of pravastatin sodium. Excipient with known effect: lactose. For the full list of excipients, see section 6.1. 3. Pharmaceutical Form Tablet. 10 mg: Yellow coloured, rounded, rectangular shaped, biconvex uncoated tablet debossed with ‘PDT’ on one side and ‘10’ on the other side. 20 mg: Yellow coloured, rounded, rectangular shaped, biconvex uncoated tablet debossed with ‘PDT’ on one side and ‘20’ on the other side. 40 mg: Yellow coloured, rounded, rectangular shaped, biconvex uncoated tablet debossed with ‘PDT’ on one side and ‘40’ on the other side. 4. Clinical Particulars 4.1 Therapeutic indications • In hypercholesterolaemic patients without clinically evident coronary heart disease, Pravastatin Mylan is indicated as an adjunct to diet to reduce the risk of fatal and non-fatal myocardial infarction, need for myocardial revascularisation procedures, and to improve survival by reducing cardiovascular deaths. • Pravastatin Mylan is indicated for the reduction of elevated total and LDL-cholesterol levels in patients with primary hypercholesterolaemia when the response to diet and other non- pharmacological measures alone have been inadequate. • Pravastatin Mylan is indicated as an adjunct to diet to slow the progressive course of atherosclerosis and reduce the incidence of clinical cardiovascular events in hypercholesterolaemic men under 75 years of age with coronary artery disease. • Coronary Artery Disease: In patients with a history of either a myocardial infarction or unstable angina pectoris, Pravastatin Mylan is indicated to reduce the risk for total mortality, CHD death, recurrent coronary event (including myocardial infarction), need for myocardial revascularisation procedures, and need for hospitalisation. -

PCSK9 Inhibition in Familial Hypercholesterolemia: a Revolution in Treatment
PCSK9 inhibition in familial hypercholesterolemia: A revolution in treatment Frederick Raal FCP(SA), FRCP, FCRPC, Cert Endo, MMED, PHD Head, Division of Endocrinology & Metabolism Director, Carbohydrate and Lipid Metabolism Research Unit Faculty of Health Sciences, University of the Witwatersrand, Johannesburg, South Africa A case of severe hypercholesterolemia Mrs. TDK 50 year old female. Well. Non smoker and no other risk factors for cardiovascular disease. Serum cholesterol measured because of positive family history of coronary artery disease, her father having undergone coronary bypass surgery at age 52. On examination she had arcus cornealis as well as marked thickening of her tendo-Achilles. Mrs TDK, a case of severe hypercholesterolemia Fasting lipid profile Total cholesterol: 15.6 mmol/L Triglycerides: 1.34 mmol/L HDL-cholesterol: 1.8 mmol/L LDL-cholesterol 13.2 mmol/L Heterozygous vs Homozygous FH Heterozygous FH Homozygous FH Prevalence 1:200- 500 Prevalence 1:300 000 -1000 000 LDL-cholesterol LDL-cholesterol 5.0-12 mmol/L 12-20 mmol/L (190-500mg/dL) (500–1000 mg/dL) CHD onset usually age 30-60 yrs CHD onset in early childhood Most patients respond to drug therapy, but individual response Poorly responsive to quite variable lipid-lowering drug therapy Nordestgaard B.G. et al. Eur Heart J 2013;34:3478-3490 Diagnostic definition of homozygous familial hypercholesterolemia . Genetic confirmation of 2 mutant alleles at the LDL receptor, APOB, PCSK9, or ARH adaptor protein gene locus OR . An untreated LDL cholesterol of 13 mmol/L (>500 mg/dL) or treated LDL cholesterol 7.8 mmol/L (≥300 mg/dL) or treated non-HDL cholesterol 8.5 mmol/L (≥330 mg/dL) together with either: - Cutaneous or tendonous xanthoma before age 10 years OR - Elevated LDL cholesterol levels before lipid-lowering therapy consistent with heterozygous FH in both parents* * Except in the case of ARH Raal FJ, Santos RD. -

Quasi-Experimental Health Policy Research: Evaluation of Universal Health Insurance and Methods for Comparative Effectiveness Research
Quasi-Experimental Health Policy Research: Evaluation of Universal Health Insurance and Methods for Comparative Effectiveness Research The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters Citation Garabedian, Laura Faden. 2013. Quasi-Experimental Health Policy Research: Evaluation of Universal Health Insurance and Methods for Comparative Effectiveness Research. Doctoral dissertation, Harvard University. Citable link http://nrs.harvard.edu/urn-3:HUL.InstRepos:11156786 Terms of Use This article was downloaded from Harvard University’s DASH repository, and is made available under the terms and conditions applicable to Other Posted Material, as set forth at http:// nrs.harvard.edu/urn-3:HUL.InstRepos:dash.current.terms-of- use#LAA Quasi-Experimental Health Policy Research: Evaluation of Universal Health Insurance and Methods for Comparative Effectiveness Research A dissertation presented by Laura Faden Garabedian to The Committee on Higher Degrees in Health Policy in partial fulfillment of the requirements for the degree of Doctor of Philosophy in the subject of Health Policy Harvard University Cambridge, Massachusetts March 2013 © 2013 – Laura Faden Garabedian All rights reserved. Professor Stephen Soumerai Laura Faden Garabedian Quasi-Experimental Health Policy Research: Evaluation of Universal Health Insurance and Methods for Comparative Effectiveness Research Abstract This dissertation consists of two empirical papers and one methods paper. The first two papers use quasi-experimental methods to evaluate the impact of universal health insurance reform in Massachusetts (MA) and Thailand and the third paper evaluates the validity of a quasi- experimental method used in comparative effectiveness research (CER). My first paper uses interrupted time series with data from IMS Health to evaluate the impact of Thailand’s universal health insurance and physician payment reform on utilization of medicines for three non-communicable diseases: cancer, cardiovascular disease and diabetes. -

Colesevelam Hydrochloride (Cholestagel) a New, Potent Bile Acid Sequestrant Associated with a Low Incidence of Gastrointestinal Side Effects
ORIGINAL INVESTIGATION Colesevelam Hydrochloride (Cholestagel) A New, Potent Bile Acid Sequestrant Associated With a Low Incidence of Gastrointestinal Side Effects Michael H. Davidson, MD; Maureen A. Dillon; Bruce Gordon, MD; Peter Jones, MD; Julie Samuels, MD; Stuart Weiss, MD; Jonathon Isaacsohn, MD; Phillip Toth, MD; Steven K. Burke, MD Objectives: To compare colesevelam hydrochloride mg/dL) (19.1%) in the 3.75-g/d colesevelam treatment (Cholestagel), a nonabsorbed hydrogel with bile acid– group. Low-density lipoprotein cholesterol concentra- sequestering properties, with placebo for its lipid- tions at the end of treatment were significantly reduced lowering efficacy, its effects on laboratory and clinical from baseline levels in the 3.0- and 3.75-g/d colesevelam safety parameters, and the incidence of adverse events. treatment groups (P = .01 and P,.001, respectively). To- tal cholesterol levels demonstrated a similar response to Methods: Following diet and placebo lead-in periods, colesevelam treatment, with an 8.1% decrease from base- placebo or colesevelam was administered at 4 dosages (1.5, line in the 3.75-g/d treatment group (P<.001). High- 2.25, 3.0, or 3.75 g/d) for 6 weeks with morning and density lipoprotein cholesterol levels rose significantly evening meals to men and women with hypercholester- in the 3.0- and 3.75-g/d colesevelam treatment groups, olemia (low-density lipoprotein cholesterol level .4.14 by 11.2% (P = .006) and 8.1% (P = .02), respectively. mmol/L [.160 mg/dL]). Patients returned to the clinic Median triglyceride levels did not change from baseline, every 2 weeks throughout the treatment period for lipid nor were there any significant differences between parameter measurements and adverse event assess- treatment groups. -

06. Barter Evolving Concepots PACE.Pdf
Evolving concepts in management of patients at increased CV risk Philip Barter, MD Sydney, Australia Evolving concepts in management of patients at increased CV risk? Philip Barter School of Medical Sciences University of New South Wales Sydney, Australia Disclosures Received honorariums for participating as a consultant or as a member of advisory boards for AMGEN, AstraZeneca, CSL-Behring, Lilly, Merck, Novartis, Pfizer and Sanofi and for giving lectures for AMGEN, AstraZeneca, Merck and Pfizer. Major risk factors for Atherosclerotic Cardiovascular Disease (ASCVD) • Age • Gender • Smoking • Elevated LDL-C • Elevated triglyceride-rich lipoproteins • Reduced HDL-C • Elevated blood pressure • Diabetes • Abdominal obesity Modifiable risk factors for ASCVD • Smoking • Elevated LDL-C • Elevated triglyceride-rich lipoproteins • Reduced HDL-C • Elevated blood pressure • Diabetes • Abdominal obesity Modifiable risk factors for ASCVD • Smoking • Elevated LDL-C • Elevated triglyceride-rich lipoproteins • Reduced HDL-C • Elevated blood pressure • Diabetes • Abdominal obesity Treatment with statins has been shown in many trials to reduce the risk of having an atherosclerotic cardiovascular event In these statin trials, the more the LDL-C is reduced, the greater is the reduction in risk of having an event. Relationship of CVD events to LDL-C reduction achieved in statin clinical trials CTT Collaboration. Lancet 2005; 366:1267-78; Lancet 2010;376:1670-81. And the lower the achieved level of LDL-C, the lower the risk of having an event Secondary Prevention -
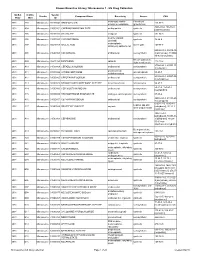
Known Bioactive Library: Microsource 1 - US Drug Collection
Known Bioactive Library: Microsource 1 - US Drug Collection ICCB-L ICCB-L Vendor Vendor Compound Name Bioactivity Source CAS Plate Well ID antifungal, inhibits Penicillium 2091 A03 Microsource 00200046 GRISEOFULVIN 126-07-8 mitosis in metaphase griseofulvum 3505-38-2, 486-16-8 2091 A04 Microsource 01500161 CARBINOXAMINE MALEATE antihistaminic synthetic [carbinoxamine] 2091 A05 Microsource 00200331 SALSALATE analgesic synthetic 552-94-3 muscle relaxant 2091 A06 Microsource 01500162 CARISOPRODOL synthetic 78-44-4 (skeletal) antineoplastic, 2091 A07 Microsource 00210369 GALLIC ACID insect galls 149-91-7 astringent, antibacterial 66592-87-8, 50370-12- 2091 A08 Microsource 01500163 CEFADROXIL antibacterial semisynthetic 2 [anhydrous], 119922- 89-9 [hemihydrate] Rheum palmatum, 2091 A09 Microsource 00211468 DANTHRON cathartic 117-10-2 Xyris semifuscata 27164-46-1, 25953-19- 2091 A10 Microsource 01500164 CEFAZOLIN SODIUM antibacterial semisynthetic 9 [cefazolin] glucocorticoid, 2091 A11 Microsource 00300024 HYDROCORTISONE adrenal glands 50-23-7 antiinflammatory 64485-93-4, 63527-52- 2091 A12 Microsource 01500165 CEFOTAXIME SODIUM antibacterial semisynthetic 6 [cefotaxime] 2091 A13 Microsource 00300029 DESOXYCORTICOSTERONE ACETATE mineralocorticoid adrenocortex 56-47-3 58-71-9, 153-61-7 2091 A14 Microsource 01500166 CEPHALOTHIN SODIUM antibacterial semisynthetic [cephalothin] 2091 A15 Microsource 00300034 TESTOSTERONE PROPIONATE androgen, antineoplastic semisynthetic 57-85-2 24356-60-3, 21593-23- 2091 A16 Microsource 01500167 CEPHAPIRIN SODIUM -

Anatomical Classification Guidelines V2020 EPHMRA ANATOMICAL
EPHMRA ANATOMICAL CLASSIFICATION GUIDELINES 2020 Anatomical Classification Guidelines V2020 "The Anatomical Classification of Pharmaceutical Products has been developed and maintained by the European Pharmaceutical Marketing Research Association (EphMRA) and is therefore the intellectual property of this Association. EphMRA's Classification Committee prepares the guidelines for this classification system and takes care for new entries, changes and improvements in consultation with the product's manufacturer. The contents of the Anatomical Classification of Pharmaceutical Products remain the copyright to EphMRA. Permission for use need not be sought and no fee is required. We would appreciate, however, the acknowledgement of EphMRA Copyright in publications etc. Users of this classification system should keep in mind that Pharmaceutical markets can be segmented according to numerous criteria." © EphMRA 2020 Anatomical Classification Guidelines V2020 CONTENTS PAGE INTRODUCTION A ALIMENTARY TRACT AND METABOLISM 1 B BLOOD AND BLOOD FORMING ORGANS 28 C CARDIOVASCULAR SYSTEM 35 D DERMATOLOGICALS 50 G GENITO-URINARY SYSTEM AND SEX HORMONES 57 H SYSTEMIC HORMONAL PREPARATIONS (EXCLUDING SEX HORMONES) 65 J GENERAL ANTI-INFECTIVES SYSTEMIC 69 K HOSPITAL SOLUTIONS 84 L ANTINEOPLASTIC AND IMMUNOMODULATING AGENTS 92 M MUSCULO-SKELETAL SYSTEM 102 N NERVOUS SYSTEM 107 P PARASITOLOGY 118 R RESPIRATORY SYSTEM 120 S SENSORY ORGANS 132 T DIAGNOSTIC AGENTS 139 V VARIOUS 141 Anatomical Classification Guidelines V2020 INTRODUCTION The Anatomical Classification was initiated in 1971 by EphMRA. It has been developed jointly by Intellus/PBIRG and EphMRA. It is a subjective method of grouping certain pharmaceutical products and does not represent any particular market, as would be the case with any other classification system. -

Medicines for Cardiovascular Health
Appendix Appendix tables Table A1: Supply of cardiovascular medicines in selected OECD countries, 2004 Medicine class Country ARA CCB BBA DIU AHYP ART GLY CHOL DDD/1,000 population/day Australia 146.9 77.0 25.5 39.4 5.2 2.2 5.3 161.8 Belgium 91.3 40.4 66.6 43.7 5.4 8.0 5.0 104.1 Czech Republic 105.1 69.9 71.3 95.5 12.2 5.6 7.1 65.0 Denmark(a) 85.1 46.2 29.9 111.5 2.7 1.6 6.3 67.2 Finland(a) 125.3 48.3 68.1 61.9 1.7 1.8 6.6 81.7 Germany(b) 143.8 48.8 67.6 65.9 11.6 2.3 10.1 57.3 Greece 149.4 68.6 30.7 35.7 6.7 5.5 9.9 84.0 Iceland 80.9 31.5 46.4 67.3 1.4 3.3 3.2 75.2 Norway(a) 99.1 46.6 39.4 45.4 n/a 1.3 4.4 110.0 Portugal(c) 109.7 37.0 18.3 42.4 0.3 7.3 6.8 71.1 Slovak Republic 88.8 63.3 42.8 31.7 10.7 4.3 6.6 31.5 Sweden(a) 85.6 40.7 54.7 87.3 1.9 1.1 6.5 75.2 ARA agents acting on renin–angiotensin system CCB calcium-channel blockers BBA beta-blocking agents DIU diuretics AHYP antihypertensives ART antiarrhythmics GLY cardiac glycosides CHOL serum-lipid-reducing agents n/a not available (a) Data for Denmark, Finland, Norway and Sweden cover supply of medicines in the community and in hospitals. -

Common Study Protocol for Observational Database Studies WP5 – Analytic Database Studies
Arrhythmogenic potential of drugs FP7-HEALTH-241679 http://www.aritmo-project.org/ Common Study Protocol for Observational Database Studies WP5 – Analytic Database Studies V 1.3 Draft Lead beneficiary: EMC Date: 03/01/2010 Nature: Report Dissemination level: D5.2 Report on Common Study Protocol for Observational Database Studies WP5: Conduct of Additional Observational Security: Studies. Author(s): Gianluca Trifiro’ (EMC), Giampiero Version: v1.1– 2/85 Mazzaglia (F-SIMG) Draft TABLE OF CONTENTS DOCUMENT INFOOMATION AND HISTORY ...........................................................................4 DEFINITIONS .................................................... ERRORE. IL SEGNALIBRO NON È DEFINITO. ABBREVIATIONS ......................................................................................................................6 1. BACKGROUND .................................................................................................................7 2. STUDY OBJECTIVES................................ ERRORE. IL SEGNALIBRO NON È DEFINITO. 3. METHODS ..........................................................................................................................8 3.1.STUDY DESIGN ....................................................................................................................8 3.2.DATA SOURCES ..................................................................................................................9 3.2.1. IPCI Database .....................................................................................................9 -

Lipotropics, Statins Therapeutic Class Review
Lipotropics, Statins Therapeutic Class Review (TCR) October 2, 2020 No part of this publication may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, digital scanning, or via any information storage or retrieval system without the express written consent of Magellan Rx Management. All requests for permission should be mailed to: Magellan Rx Management Attention: Legal Department 6950 Columbia Gateway Drive Columbia, Maryland 21046 The materials contained herein represent the opinions of the collective authors and editors and should not be construed to be the official representation of any professional organization or group, any state Pharmacy and Therapeutics committee, any state Medicaid Agency, or any other clinical committee. This material is not intended to be relied upon as medical advice for specific medical cases and nothing contained herein should be relied upon by any patient, medical professional or layperson seeking information about a specific course of treatment for a specific medical condition. All readers of this material are responsible for independently obtaining medical advice and guidance from their own physician and/or other medical professional in regard to the best course of treatment for their specific medical condition. This publication, inclusive of all forms contained herein, is intended to be educational in nature and is intended to be used for informational purposes only. Send comments and suggestions to [email protected]. October -

Probucol Inhibits the Initiation of Atherosclerosis in Cholesterol-Fed
Niimi et al. Lipids in Health and Disease 2013, 12:166 http://www.lipidworld.com/content/12/1/166 RESEARCH Open Access Probucol inhibits the initiation of atherosclerosis in cholesterol-fed rabbits Manabu Niimi1, Yuka Keyamura2, Masanori Nozako2, Takashi Koyama2, Masayuki Kohashi2, Reiko Yasufuku2, Tomohiro Yoshikawa2 and Jianglin Fan1* Abstract Background: Probucol and statin are often prescribed for treating atherosclerosis. These two drugs exhibit different mechanisms but it is unknown whether they have the same anti-atherogenic properties. In the current study, we examined whether these two drugs at optimal doses could inhibit the initiation of atherosclerosis in cholesterol-fed rabbits in the same way. Methods: New Zealand White rabbits were fed a cholesterol-rich diet for 5 weeks to produce the early-stage lesions of atherosclerosis. Drug-treated rabbits were administered either probucol or atorvastatin and serum lipids and aortic atherosclerotic lesions were compared with those in a control group. Results: Atorvastatin treatment significantly reduced serum total cholesterol levels while probucol treatment led to significant reduction of high-density lipoprotein cholesterol levels without changing total cholesterol levels compared with those in the control group. Compared with the control, probucol treatment led to 65% (p < 0.01) reduction while atorvastatin treatment led to 23% (p = 0.426) reduction of the aortic lesion area. Histological and immunohistochemical analyses revealed that the lesions of the probucol-treated group were characterized by remarkable reduction of monocyte adherence to endothelial cells and macrophage accumulation in the intima compared with those of both atorvastatin and control groups. Furthermore, low-density lipoprotein (LDL) isolated from the probucol group exhibited prominent anti-oxidative reaction, which was not present in LDL isolated from either the atorvastatin-treated or the control group.