Paradoxical Enhancement of Atherosclerosis by Probucol Treatment in Apolipoprotein Eðdeficient Mice
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Deliverable 5.A Interim Report on the Study Results APPENDIX 2
Deliverable 5.a Interim report on the study results APPENDIX 2: Algorithms used to identify study variables for service contract EMA/2011/38/CN ‐ PIOGLITAZONE November 28th 2012 D5.a Interim report on the study results (Appendix 2) for Service Contract EMA/2011/38/CN PIOGLITAZONE Author(s): Vera Ehrenstein (AUH‐AS) APPENDIX 2. ALGORITHMS USED TO IDENTIFY STUDY VARIABLES Algorithms for AU Database DISEASE/CONDITION ICD-8 CODE (1977-1993) ICD-10 CODE (1994-) Diabetes type 2 250.00; 250.06; 250.07; 250.09 E11.0; E11.1; E11.9 Cancer of bladder 188 C67 Haematuria N/A R31 Haematuria, unspecified B18, K70.0–K70.3, K70.9, K71, K73, Mild hepatic impairment 571, 573.01, 573.04 K74, K76.0 Moderate to severe hepatic 070.00, 070.02, 070.04, 070.06, B15.0, B16.0, B16.2, B19.0, K70.4, impairment 070.08, 573.00, 456.00–456.09 K72, K76.6, I85 Acute myocardial infarction 410 I21-I23 Acute coronary syndrome 410, 413 I20-I24 Ischemic heart disease 410-414 I20-I25 427.09, 427.10, 427.11, 427.19, Congestive heart failure I50, I11.0, I13.0,I13.2 428.99, 782.49; Acute renal failure N/A N17 Diabetic coma N/A E10.0, E11.0, E12.0,E13.0, E14.0 Diabetic acidosis N/A E10.1, E11.1, E12.1,E13.1, E14.1 F10.1-F10.9, G31.2, G62.1, G72.1, Alcoholism 291, 303, 577.10, 571.09, 571.10 I42.6, K29.2, K86.0, Z72.1 Obesity 277.99 E65-E66 D5.a Interim report on the study results (Appendix 2) for Service Contract EMA/2011/38/CN PIOGLITAZONE Author(s): Vera Ehrenstein (AUH‐AS) Algorithms for defining acute events in Denmark, ICD-10 code Event ICD-10 code I21.x, I23.x http://apps.who.int/classifications/icd10/browse/2010/en#/I21 -

Hypertension, Cardiac Hypertrophy, and Sudden Death in Mice Lacking
Proc. Natl. Acad. Sci. USA Vol. 94, pp. 14730–14735, December 1997 Medical Sciences Hypertension, cardiac hypertrophy, and sudden death in mice lacking natriuretic peptide receptor A (gene targetingyechocardiographyycardiac dilatationyinterstitial fibrosisyaortic dissection) PAULA M. OLIVER*, JENNIFER E. FOX*, RON KIM*, HOWARD A. ROCKMAN†,HYUNG-SUK KIM*, i ROBERT L. REDDICK‡,KAILASH N. PANDEY§,SHARON L. MILGRAM¶,OLIVER SMITHIES*, AND NOBUYO MAEDA* *Department of Pathology, University of North Carolina, 710 Brinkhous-Bullitt Building, Chapel Hill, NC 27599-7525; Departments of †Medicine and ¶Physiology, University of North Carolina, Chapel Hill, NC 27599; ‡Department of Pathology, The University of Texas Health Science Center, 7703 Floyd Curl Drive, San Antonio, TX 78284; and §Department of Biochemistry and Molecular Biology, Medical College of Georgia, Augusta, GA 30912 Contributed by Oliver Smithies, October 27, 1997 ABSTRACT Natriuretic peptides, produced in the heart, of blood pressure and in the cardiovascular response to bind to the natriuretic peptide receptor A (NPRA) and cause sustained hypertension we have made mice completely lacking vasodilation and natriuresis important in the regulation of this receptor. Our results demonstrate that NPRA deficiency blood pressure. We here report that mice lacking a functional in mice leads to elevated blood pressures and, particularly in Npr1 gene coding for NPRA have elevated blood pressures and males, to cardiac hypertrophy and sudden death. hearts exhibiting marked hypertrophy with interstitial fibro- sis resembling that seen in human hypertensive heart disease. Echocardiographic evaluation of the mice demonstrated a MATERIALS AND METHODS compensated state of systemic hypertension in which cardiac Gene Targeting. Three electroporations of embryonic stem hypertrophy and dilatation are evident but with no reduction (ES) cells (E14TG2a) were carried out as described (11). -

Consumer Medicine Information
NEW ZEALAND DATA SHEET PRAVASTATIN MYLAN 1. Product Name Pravastatin Mylan, 10 mg, 20 mg and 40 mg, tablets 2. Qualitative and Quantitative Composition Each tablet contains either 10 mg, 20 mg or 40 mg of pravastatin sodium. Excipient with known effect: lactose. For the full list of excipients, see section 6.1. 3. Pharmaceutical Form Tablet. 10 mg: Yellow coloured, rounded, rectangular shaped, biconvex uncoated tablet debossed with ‘PDT’ on one side and ‘10’ on the other side. 20 mg: Yellow coloured, rounded, rectangular shaped, biconvex uncoated tablet debossed with ‘PDT’ on one side and ‘20’ on the other side. 40 mg: Yellow coloured, rounded, rectangular shaped, biconvex uncoated tablet debossed with ‘PDT’ on one side and ‘40’ on the other side. 4. Clinical Particulars 4.1 Therapeutic indications • In hypercholesterolaemic patients without clinically evident coronary heart disease, Pravastatin Mylan is indicated as an adjunct to diet to reduce the risk of fatal and non-fatal myocardial infarction, need for myocardial revascularisation procedures, and to improve survival by reducing cardiovascular deaths. • Pravastatin Mylan is indicated for the reduction of elevated total and LDL-cholesterol levels in patients with primary hypercholesterolaemia when the response to diet and other non- pharmacological measures alone have been inadequate. • Pravastatin Mylan is indicated as an adjunct to diet to slow the progressive course of atherosclerosis and reduce the incidence of clinical cardiovascular events in hypercholesterolaemic men under 75 years of age with coronary artery disease. • Coronary Artery Disease: In patients with a history of either a myocardial infarction or unstable angina pectoris, Pravastatin Mylan is indicated to reduce the risk for total mortality, CHD death, recurrent coronary event (including myocardial infarction), need for myocardial revascularisation procedures, and need for hospitalisation. -

PCSK9 Inhibition in Familial Hypercholesterolemia: a Revolution in Treatment
PCSK9 inhibition in familial hypercholesterolemia: A revolution in treatment Frederick Raal FCP(SA), FRCP, FCRPC, Cert Endo, MMED, PHD Head, Division of Endocrinology & Metabolism Director, Carbohydrate and Lipid Metabolism Research Unit Faculty of Health Sciences, University of the Witwatersrand, Johannesburg, South Africa A case of severe hypercholesterolemia Mrs. TDK 50 year old female. Well. Non smoker and no other risk factors for cardiovascular disease. Serum cholesterol measured because of positive family history of coronary artery disease, her father having undergone coronary bypass surgery at age 52. On examination she had arcus cornealis as well as marked thickening of her tendo-Achilles. Mrs TDK, a case of severe hypercholesterolemia Fasting lipid profile Total cholesterol: 15.6 mmol/L Triglycerides: 1.34 mmol/L HDL-cholesterol: 1.8 mmol/L LDL-cholesterol 13.2 mmol/L Heterozygous vs Homozygous FH Heterozygous FH Homozygous FH Prevalence 1:200- 500 Prevalence 1:300 000 -1000 000 LDL-cholesterol LDL-cholesterol 5.0-12 mmol/L 12-20 mmol/L (190-500mg/dL) (500–1000 mg/dL) CHD onset usually age 30-60 yrs CHD onset in early childhood Most patients respond to drug therapy, but individual response Poorly responsive to quite variable lipid-lowering drug therapy Nordestgaard B.G. et al. Eur Heart J 2013;34:3478-3490 Diagnostic definition of homozygous familial hypercholesterolemia . Genetic confirmation of 2 mutant alleles at the LDL receptor, APOB, PCSK9, or ARH adaptor protein gene locus OR . An untreated LDL cholesterol of 13 mmol/L (>500 mg/dL) or treated LDL cholesterol 7.8 mmol/L (≥300 mg/dL) or treated non-HDL cholesterol 8.5 mmol/L (≥330 mg/dL) together with either: - Cutaneous or tendonous xanthoma before age 10 years OR - Elevated LDL cholesterol levels before lipid-lowering therapy consistent with heterozygous FH in both parents* * Except in the case of ARH Raal FJ, Santos RD. -

Quasi-Experimental Health Policy Research: Evaluation of Universal Health Insurance and Methods for Comparative Effectiveness Research
Quasi-Experimental Health Policy Research: Evaluation of Universal Health Insurance and Methods for Comparative Effectiveness Research The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters Citation Garabedian, Laura Faden. 2013. Quasi-Experimental Health Policy Research: Evaluation of Universal Health Insurance and Methods for Comparative Effectiveness Research. Doctoral dissertation, Harvard University. Citable link http://nrs.harvard.edu/urn-3:HUL.InstRepos:11156786 Terms of Use This article was downloaded from Harvard University’s DASH repository, and is made available under the terms and conditions applicable to Other Posted Material, as set forth at http:// nrs.harvard.edu/urn-3:HUL.InstRepos:dash.current.terms-of- use#LAA Quasi-Experimental Health Policy Research: Evaluation of Universal Health Insurance and Methods for Comparative Effectiveness Research A dissertation presented by Laura Faden Garabedian to The Committee on Higher Degrees in Health Policy in partial fulfillment of the requirements for the degree of Doctor of Philosophy in the subject of Health Policy Harvard University Cambridge, Massachusetts March 2013 © 2013 – Laura Faden Garabedian All rights reserved. Professor Stephen Soumerai Laura Faden Garabedian Quasi-Experimental Health Policy Research: Evaluation of Universal Health Insurance and Methods for Comparative Effectiveness Research Abstract This dissertation consists of two empirical papers and one methods paper. The first two papers use quasi-experimental methods to evaluate the impact of universal health insurance reform in Massachusetts (MA) and Thailand and the third paper evaluates the validity of a quasi- experimental method used in comparative effectiveness research (CER). My first paper uses interrupted time series with data from IMS Health to evaluate the impact of Thailand’s universal health insurance and physician payment reform on utilization of medicines for three non-communicable diseases: cancer, cardiovascular disease and diabetes. -

Apolipoprotein E Isoforms in Diabetic Dyslipidemia and Atherosclerosis
Apolipoprotein E isoforms in diabetic dyslipidemia and atherosclerosis Lance Johnson Diabetic Dyslipidemia Cluster of harmful changes to lipoprotein metabolism commonly noted in diabetic patients Characterized by high VLDL triglycerides, low HDL-cholesterol, and a increase in small, dense LDL particles Increases the risk of developing atherosclerosis Apolipoprotein E Isoforms ApoE2 ApoE3 ApoE4 position 112 cys cys arg position 158 cys arg arg frequency 7.2% 78.3% 14.3% LDLR affinity <1% 100% >100% plasma TC -6% ref. +6% MI risk reduced ref. Increased (type III) Plasma lipoproteins Normal LDLR Increased LDLR 2m (350mg/dl) 2h (100mg/dl) 60 VLDL HDL Mice with 40 On a HFW LDL Human apoE2 HDL diet Atherosclerosis No plaques 20 (poor ligand) Cholesterol VLDL LDL 0 10 20 30 10 20 30 60 3m (100mg/dl) 3h (60mg/dl) Mice with HDL 40 Human apoE3 No plaques HDL No plaques (reference) 20 VLDL Cholesterol VLDL 0 10 20 30 10 20 30 4m (100mg/dl) 4h (250mg/dl) 60 HDL VLDL Mice with 40 Human apoE4 No plaques Atherosclerosis HDL 20 (better ligand) Cholesterol VLDL 0 10 20 30 10 20 30 ApoE -Trapping by LDL receptor Figure 2 ApoE trapping. Efficient transfer of apoE4 facilitates internalization of large lipoproteins in 4m mice. In 4h mice, the efficiency of transfer is reduced because apoE4 are trapped by the increased LDLR. Akita-diabetic mice with human apoE isoforms Survival With normal LDLR With human LDLR Diabetic Human ApoE Mice: Survival Rate 110 100 4mAE4 3hA 90 80 3mAE3 2hA 70 60 50 40 30 2mAE2 Percent survival 4hA 20 10 0 0 1 2 3 4 5 6 7 Months of age 800 600 400 Glucose 200 0 2m 2h 3m 3h 4m 4h 200 2mAkita 4h-Akita 150 100 Cholesterol 50 0 2m 2h 3m 3h 4m 4h 150 100 50 Triglycerides 0 2m 2h 3m 3h 4m 4h Two of two -6mo old 4hAkita mice on a low cholesterol diet had atherosclerosis despite of low plasma cholesterol (<120 mg/dl) levels This is the first mouse model of atherosclerosis for which diabetes is required. -

Colesevelam Hydrochloride (Cholestagel) a New, Potent Bile Acid Sequestrant Associated with a Low Incidence of Gastrointestinal Side Effects
ORIGINAL INVESTIGATION Colesevelam Hydrochloride (Cholestagel) A New, Potent Bile Acid Sequestrant Associated With a Low Incidence of Gastrointestinal Side Effects Michael H. Davidson, MD; Maureen A. Dillon; Bruce Gordon, MD; Peter Jones, MD; Julie Samuels, MD; Stuart Weiss, MD; Jonathon Isaacsohn, MD; Phillip Toth, MD; Steven K. Burke, MD Objectives: To compare colesevelam hydrochloride mg/dL) (19.1%) in the 3.75-g/d colesevelam treatment (Cholestagel), a nonabsorbed hydrogel with bile acid– group. Low-density lipoprotein cholesterol concentra- sequestering properties, with placebo for its lipid- tions at the end of treatment were significantly reduced lowering efficacy, its effects on laboratory and clinical from baseline levels in the 3.0- and 3.75-g/d colesevelam safety parameters, and the incidence of adverse events. treatment groups (P = .01 and P,.001, respectively). To- tal cholesterol levels demonstrated a similar response to Methods: Following diet and placebo lead-in periods, colesevelam treatment, with an 8.1% decrease from base- placebo or colesevelam was administered at 4 dosages (1.5, line in the 3.75-g/d treatment group (P<.001). High- 2.25, 3.0, or 3.75 g/d) for 6 weeks with morning and density lipoprotein cholesterol levels rose significantly evening meals to men and women with hypercholester- in the 3.0- and 3.75-g/d colesevelam treatment groups, olemia (low-density lipoprotein cholesterol level .4.14 by 11.2% (P = .006) and 8.1% (P = .02), respectively. mmol/L [.160 mg/dL]). Patients returned to the clinic Median triglyceride levels did not change from baseline, every 2 weeks throughout the treatment period for lipid nor were there any significant differences between parameter measurements and adverse event assess- treatment groups. -

06. Barter Evolving Concepots PACE.Pdf
Evolving concepts in management of patients at increased CV risk Philip Barter, MD Sydney, Australia Evolving concepts in management of patients at increased CV risk? Philip Barter School of Medical Sciences University of New South Wales Sydney, Australia Disclosures Received honorariums for participating as a consultant or as a member of advisory boards for AMGEN, AstraZeneca, CSL-Behring, Lilly, Merck, Novartis, Pfizer and Sanofi and for giving lectures for AMGEN, AstraZeneca, Merck and Pfizer. Major risk factors for Atherosclerotic Cardiovascular Disease (ASCVD) • Age • Gender • Smoking • Elevated LDL-C • Elevated triglyceride-rich lipoproteins • Reduced HDL-C • Elevated blood pressure • Diabetes • Abdominal obesity Modifiable risk factors for ASCVD • Smoking • Elevated LDL-C • Elevated triglyceride-rich lipoproteins • Reduced HDL-C • Elevated blood pressure • Diabetes • Abdominal obesity Modifiable risk factors for ASCVD • Smoking • Elevated LDL-C • Elevated triglyceride-rich lipoproteins • Reduced HDL-C • Elevated blood pressure • Diabetes • Abdominal obesity Treatment with statins has been shown in many trials to reduce the risk of having an atherosclerotic cardiovascular event In these statin trials, the more the LDL-C is reduced, the greater is the reduction in risk of having an event. Relationship of CVD events to LDL-C reduction achieved in statin clinical trials CTT Collaboration. Lancet 2005; 366:1267-78; Lancet 2010;376:1670-81. And the lower the achieved level of LDL-C, the lower the risk of having an event Secondary Prevention -

Original Article -Lipoic Acid Prevents the Increase in Atherosclerosis Induced By
Original Article ␣-Lipoic Acid Prevents the Increase in Atherosclerosis Induced by Diabetes in Apolipoprotein E–Deficient Mice Fed High-Fat/Low-Cholesterol Diet Xianwen Yi and Nobuyo Maeda Considerable evidence indicates that hyperglycemia in- LDL and other lipids by free radicals is one of the most creases oxidative stress and contributes to the increased important factors for the initiation of atherosclerosis (5,6). incidence of atherosclerosis and cardiovascular complica- Oxidative stress also facilitates endothelial cell dysfunc- tions in diabetic patients. To examine the effect of ␣-lipoic tion (7). However, evidence linking antioxidant vitamins to acid, a potent natural antioxidant, on atherosclerosis in diabetes complications in humans is still inconclusive. For diabetic mice, 3-month-old apolipoprotein (apo) E-defi- cient (apoE؊/؊) mice were made diabetic by administering example, although treatment with a high dose of vitamin E streptozotocin (STZ). At 4 weeks after starting the STZ (1,800 IU/day) appeared to be effective in normalizing administration, a high-fat diet with or without ␣-lipoic acid retinal hemodynamic abnormalities and improving renal (1.65 g/kg) was given to the mice and to nondiabetic function in type 1 diabetes in a small randomized trial (8), apoE؊/؊ controls. At 20 weeks, markers of oxidative stress vitamin E at a dose of 400 IU/day had no effect on ؊/؊ were significantly lower in both the diabetic apoE mice cardiovascular outcomes or nephropathy in high-risk pa- ␣ ؊/؊ and their nondiabetic apoE controls with -lipoic acid tients with diabetes, as reported in the HOPE (Heart supplement than in those without it. Remarkably, ␣-lipoic acid completely prevented the increase in plasma total Outcomes Prevention Evaluation) study and MICRO-HOPE cholesterol, atherosclerotic lesions, and the general dete- (Microalbuminuria, Cardiovascular, and Renal Outcomes- rioration of health caused by diabetes. -

Hypertension and Abnormal Fat Distribution but Not Insulin Resistance in Mice with P465L Pparγ
Hypertension and abnormal fat distribution but not insulin resistance in mice with P465L PPARγ Yau-Sheng Tsai, … , Jason K. Kim, Nobuyo Maeda J Clin Invest. 2004;114(2):240-249. https://doi.org/10.1172/JCI20964. Article Cardiology Peroxisome proliferator–activated receptor γ (PPARγ), the molecular target of a class of insulin sensitizers, regulates adipocyte differentiation and lipid metabolism. A dominant negative P467L mutation in the ligand-binding domain of PPARγ in humans is associated with severe insulin resistance and hypertension. Homozygous mice with the equivalent P465L mutation die in utero. Heterozygous mice grow normally and have normal total adipose tissue weight. However, they have reduced interscapular brown adipose tissue and intra-abdominal fat mass, and increased extra-abdominal subcutaneous fat, compared with wild-type mice. They have normal plasma glucose levels and insulin sensitivity, and increased glucose tolerance. However, during high-fat feeding, their plasma insulin levels are mildly elevated in association with a significant increase in pancreatic islet mass. They are hypertensive, and expression of the angiotensinogen gene is increased in their subcutaneous adipose tissues. The effects of P465L on blood pressure, fat distribution, and insulin sensitivity are the same in both male and female mice regardless of diet and age. Thus the P465L mutation alone is sufficient to cause abnormal fat distribution and hypertension but not insulin resistance in mice. These results provide genetic evidence for a critical role for PPARγ in blood pressure regulation that is not dependent on altered insulin sensitivity. Find the latest version: https://jci.me/20964/pdf Research article Related Commentary, page 163 Hypertension and abnormal fat distribution but not insulin resistance in mice with P465L PPARγ Yau-Sheng Tsai,1 Hyo-Jeong Kim,2 Nobuyuki Takahashi,1 Hyung-Suk Kim,1 John R. -

Professor Oliver Smithies Was Born on June 23, 1925, in Halifax, England, and As a Child Went to a Small School in Copley Village Near to Halifax
Professor Oliver Smithies was born on June 23, 1925, in Halifax, England, and as a child went to a small school in Copley village near to Halifax. Subsequently he was a student at Heath Grammar School in Halifax until he was awarded a scholarship from Balliol College, Oxford University. At Oxford he received a Bachelor of Arts Degree in Physiology with First Class Honors in 1946. In 1951, he obtained his M.A. and D.Phil. degrees in Biochemistry from Oxford. Professor Smithies then moved to the United States as a Postdoctoral Fellow in Physical Chemistry at the University of Wisconsin. After two years at Wisconsin, Professor Smithies accepted a position at the Connaught Medical Research Laboratories in Toronto and stayed there from 1953 to 1960, first as a Research Assistant and then as a Research Associate. In 1960, Professor Smithies returned to Wisconsin as Assistant Professor of Medical Genetics and Genetics, advancing to Full Professor by 1963. He was named the Leon J. Cole Professor in 1971. In 1988, he joined the Department of Pathology at the University of North Carolina, as the Excellence Professor of Pathology. He is now the Weatherspoon Eminent Distinguished Professor of Pathology and Laboratory Medicine at UNC, and remains an actively engaged scientist working at the bench in the laboratory he shares with his wife, Nobuyo Maeda who is the Robert H. Wagner Distinguished Professor of Pathology and Laboratory Medicine. In the mid-1950s, Professor Smithies described the first high resolution electrophoresis system (starch gel), and with it he discovered that normal humans have unsuspected inherited differences in their proteins. -
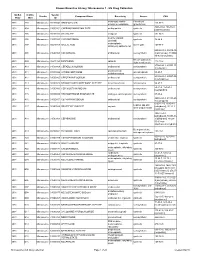
Known Bioactive Library: Microsource 1 - US Drug Collection
Known Bioactive Library: Microsource 1 - US Drug Collection ICCB-L ICCB-L Vendor Vendor Compound Name Bioactivity Source CAS Plate Well ID antifungal, inhibits Penicillium 2091 A03 Microsource 00200046 GRISEOFULVIN 126-07-8 mitosis in metaphase griseofulvum 3505-38-2, 486-16-8 2091 A04 Microsource 01500161 CARBINOXAMINE MALEATE antihistaminic synthetic [carbinoxamine] 2091 A05 Microsource 00200331 SALSALATE analgesic synthetic 552-94-3 muscle relaxant 2091 A06 Microsource 01500162 CARISOPRODOL synthetic 78-44-4 (skeletal) antineoplastic, 2091 A07 Microsource 00210369 GALLIC ACID insect galls 149-91-7 astringent, antibacterial 66592-87-8, 50370-12- 2091 A08 Microsource 01500163 CEFADROXIL antibacterial semisynthetic 2 [anhydrous], 119922- 89-9 [hemihydrate] Rheum palmatum, 2091 A09 Microsource 00211468 DANTHRON cathartic 117-10-2 Xyris semifuscata 27164-46-1, 25953-19- 2091 A10 Microsource 01500164 CEFAZOLIN SODIUM antibacterial semisynthetic 9 [cefazolin] glucocorticoid, 2091 A11 Microsource 00300024 HYDROCORTISONE adrenal glands 50-23-7 antiinflammatory 64485-93-4, 63527-52- 2091 A12 Microsource 01500165 CEFOTAXIME SODIUM antibacterial semisynthetic 6 [cefotaxime] 2091 A13 Microsource 00300029 DESOXYCORTICOSTERONE ACETATE mineralocorticoid adrenocortex 56-47-3 58-71-9, 153-61-7 2091 A14 Microsource 01500166 CEPHALOTHIN SODIUM antibacterial semisynthetic [cephalothin] 2091 A15 Microsource 00300034 TESTOSTERONE PROPIONATE androgen, antineoplastic semisynthetic 57-85-2 24356-60-3, 21593-23- 2091 A16 Microsource 01500167 CEPHAPIRIN SODIUM