A Review of Information on Temporarily Open/Closed Estuaries
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Gamtoos Estuary Management Plan Volume I Situation Assessment
Enviro-Fish Africa (Pty) Ltd. C.A.P.E. ESTUARIES MANAGEMENT PROGRAMME GAMTOOS ESTUARY MANAGEMENT PLAN VOLUME I SITUATION ASSESSMENT (STATE OF PLAY REPORT) Produced by: Enviro-Fish Africa (Pty) Ltd. P.O. Box 94 Grahamstown 6140 Produced for: Cape Nature December 2007 C.A.P.E. Estuaries Management Programme; Gamtoos Estuary Management Plan: Situation Assessment 1 Enviro-Fish Africa (Pty) Ltd. 1. INTRODUCTION Estuarine ecosystems are not isolated systems. They form an interface between marine and freshwater systems and are part of regional, national and global ecosystems either directly via water flows or indirectly through the movement of fauna. In addition to the biota that these estuaries support, they provide a range of goods and services (uses) to the inhabitants of the various regions. Disturbances in one estuary can influence a wide variety of habitats and organisms in the broader freshwater or marine ecosystem. Thus, the interaction between the systems and users creates a delicate balance, the sustainability of which needs to be addressed by some form of management plan. In order to address this balance in a consistent manner in the Cape Floristic Region (CFR), the Cape Action for People and the Environment (C.A.P.E.) Estuaries Management Programme has developed a holistic and inclusive management process representative of all stakeholders. The programme is governed by a Task Team comprising of officials from C.A.P.E., CapeNature, Marine and Coastal Management (MCM), the Department of Water Affairs and Forestry (DWAF), the Eastern Cape Parks Board (ECPB) and the Council for Scientific and Industrial Research (CSIR), which heads the technical support group. -

Water for Food and Ecosystems in the Baviaanskloof Mega Reserve Land and Water Resources Assessment in the Baviaanskloof, Easter
Water for Food and Ecosystems in the Baviaanskloof Mega Reserve Land and water resources assessment in the Baviaanskloof, Eastern Cape Province, South Africa H.C. Jansen Alterra-report 1812 Alterra, Wageningen, 2008 ABSTRACT Jansen, H.C., 2008. Walerfor bood and hicosystems in the baviaanskloofMega Reserve. IMnd and water resources assessment in the Baviaanskloof,Hastern Cape Province, South Africa. Wageningen, Alterra, Alterra-report 1812. 80 pages; 21 figs.; 6 tables.; 18 refs. This report describes the results of the land and water assessment for the project 'Water for Food and Ecosystems in the Baviaanskloof Mega Reserve'. Aim of the project is to conserve the biodiversity in a more sustainable way, by optimizing water for ecosystems, agricultural and domestic use, in a sense that its also improving rural livelihoods in the Baviaanskloof. In this report an assessment of the land and water system is presented, which forms a basis for the development and implementation of land and water policies and measures. Keywords: competing claims, IWRM, land management, nature conservation, policy support, water management, water retention ISSN 1566-7197 The pdf file is tree of charge and can he downloaded vi«i the website www.ahctra.wur.nl (go lo Alterra reports). Alterra docs not deliver printed versions ol the Altena reports. Punted versions can be ordered via the external distributor. I-or oidcrmg have a look at www.li tx> ni l) ljtl.nl/mppcirtc ilser vice . © 2008 Alterra P.O. Box 47; 6700 AA Wageningen; The Netherlands Phone: + 31 317 484700; fax: +31 317 419000; e-mail: info.alterra@,wur.nl No part of this publication may be reproduced or published in any form or by any means, or stored in a database or retrieval system without the written permission of Alterra. -

Population Trends of Seabirds Breeding in South Africa's Eastern Cape and the Possible Influence of Anthropogenic and Environ
Crawford et al.: Population trends of seabirds breeding in South Africa 159 POPULATION TRENDS OF SEABIRDS BREEDING IN SOUTH AFRICA’S EASTERN CAPE AND THE POSSIBLE INFLUENCE OF ANTHROPOGENIC AND ENVIRONMENTAL CHANGE ROBERT J.M. CRAWFORD,1,2 PHILIP A. WHITTINGTON,3,4 A. PAUL MARTIN,5 ANTHONY J. TREE4,6 & AZWIANEWI B. MAKHADO1 1Department of Environmental Affairs and Tourism, Marine and Coastal Management, Private Bag X2, Rogge Bay, 8012, South Africa ([email protected]) 2Animal Demography Unit, Department of Zoology, University of Cape Town, Rondebosch, 7701, South Africa 3East London Museum, PO Box 11021, Southernwood, 5213, South Africa 4Department of Zoology, PO Box 77000, Nelson Mandela Metropolitan University, Port Elizabeth, 6031, South Africa 5PO Box 61029, Bluewater Bay, 6212, South Africa 6PO Box 211, Bathurst, 6166, South Africa Received 28 August 2008, accepted 4 April 2009 SUMMARY CRAWFORD, R.J.M., WHITTINGTON, P.A., MARTIN, A.P., TREE, A.J. & MAKHADO, A.B. 2009. Population trends of seabirds breeding in South Africa’s Eastern Cape and the possible influence of anthropogenic and environmental change. Marine Ornithology 37: 159–174. Eleven species of seabird breed in South Africa’s Eastern Cape Province. Numbers of African Penguin Spheniscus demersus and Cape Gannet Morus capensis in the province increased in the 20th century, but penguins decreased in the early 21st century. A recent eastward displacement of Sardine Sardinops sagax off South Africa increased the availability of this food source to gannets but did not benefit penguins, which have a shorter foraging range. Fishing and harbour developments may have influenced the recent decrease of penguins. -

Misgund Orchards
MISGUND ORCHARDS ENVIRONMENTAL AUDIT 2014 Grey Rhebok Pelea capreolus Prepared for Mr Wayne Baldie By Language of the Wilderness Foundation Trust In March 2002 a baseline environmental audit was completed by Conservation Management Services. This foundational document has served its purpose. The two (2) recommendations have been addressed namely; a ‘black wattle control plan’ in conjunction with Working for Water Alien Eradication Programme and a survey of the fish within the rivers was also addressed. Furthermore updated species lists have resulted (based on observations and studies undertaken within the region). The results of these efforts have highlighted the significance of the farm Misgund Orchards and the surrounds, within the context of very special and important biodiversity. Misgund Orchards prides itself with a long history of fruit farming excellence, and has strived to ensure a healthy balance between agricultural priorities and our environment. Misgund Orchards recognises the need for a more holistic and co-operative regional approach towards our environment and needs to adapt and design a more sustainable approach. The context of Misgund Orchards is significant, straddling the protected areas Formosa Forest Reserve (Niekerksberg) and the Baviaanskloof Mega Reserve. A formidable mountain wilderness with World Heritage Status and a Global Biodiversity Hotspot (See Map 1 overleaf). Rhombic egg eater Dasypeltis scabra MISGUND ORCHARDS Langkloof Catchment MAP 1 The regional context of Misgund Orchards becomes very apparent, where the obvious strategic opportunity exists towards creating a bridge of corridors linking the two mountain ranges Tsitsikamma and Kouga (south to north). The environmental significance of this cannot be overstated – essentially creating a protected area from the ocean into the desert of the Klein-karoo, a traverse of 8 biomes, a veritable ‘garden of Eden’. -

Statistical Based Regional Flood Frequency Estimation Study For
Statistical Based Regional Flood Frequency Estimation Study for South Africa Using Systematic, Historical and Palaeoflood Data Pilot Study – Catchment Management Area 15 by D van Bladeren, P K Zawada and D Mahlangu SRK Consulting & Council for Geoscience Report to the Water Research Commission on the project “Statistical Based Regional Flood Frequency Estimation Study for South Africa using Systematic, Historical and Palaeoflood Data” WRC Report No 1260/1/07 ISBN 078-1-77005-537-7 March 2007 DISCLAIMER This report has been reviewed by the Water Research Commission (WRC) and approved for publication. Approval does not signify that the contents necessarily reflect the views and policies of the WRC, nor does mention of trade names or commercial products constitute endorsement or recommendation for use EXECUTIVE SUMMARY INTRODUCTION During the past 10 years South Africa has experienced several devastating flood events that highlighted the need for more accurate and reasonable flood estimation. The most notable events were those of 1995/96 in KwaZulu-Natal and north eastern areas, the November 1996 floods in the Southern Cape Region, the floods of February to March 2000 in the Limpopo, Mpumalanga and Eastern Cape provinces and the recent floods in March 2003 in Montagu in the Western Cape. These events emphasized the need for a standard approach to estimate flood probabilities before developments are initiated or existing developments evaluated for flood hazards. The flood peak magnitudes and probabilities of occurrence or return period required for flood lines are often overlooked, ignored or dealt with in a casual way with devastating effects. The National Disaster and new Water Act and the rapid rate at which developments are being planned will require the near mass production of flood peak probabilities across the country that should be consistent, realistic and reliable. -

Disaster Management Plan
SARAH BAARTMAN DISTRICT MUNICIPALITY DISASTER RISK MANANAGEMENT PLAN Contents Part 1 Introduction Foreword Authority for Planning Amendments Setting the scene Definitions The Municipal disaster management framework setting out Responsibilities and co-ordination Who will implement the plan and the custodian of the plan? Part 2 Description of Sarah Baartman District Part 3 Risk Management Process Section 1: Introduction Section 2: Risk Assessment Section 3: Assessing the Local Risk Section 4: Summarised Risk Metrics Section 5: Top Priority Risk Assessment Summaries Section 6: Impact of relevant global risk conditions Section 7: Recommended way forward Section 8: Generic Municipal Information Part 5 External Liaison and Stakeholders Participation Part 6 Operational Planning Part 7 Disaster Management SOPs (standard operating Protocols) PART ONE Foreword “Whilst hazards are inevitable, and the elimination of all risks impossible, there are many technical measures, traditional practices and public experiences that can reduce the extent or severity of economic, environmental and social disasters.” South Africa is susceptible to a range of hazards, both natural and technological, which have the capacity to impact significantly on national and municipal development strategies and initiatives. The sustainability of such initiatives will therefore depend very much on how successfully risks associated with these hazards can be managed. This Disaster Management Plan has been developed by Sarah Baartman District Municipality Disaster Management Centre, -

Explore the Eastern Cape Province
Cultural Guiding - Explore The Eastern Cape Province Former President Nelson Mandela, who was born and raised in the Transkei, once said: "After having travelled to many distant places, I still find the Eastern Cape to be a region full of rich, unused potential." 2 – WildlifeCampus Cultural Guiding Course – Eastern Cape Module # 1 - Province Overview Component # 1 - Eastern Cape Province Overview Module # 2 - Cultural Overview Component # 1 - Eastern Cape Cultural Overview Module # 3 - Historical Overview Component # 1 - Eastern Cape Historical Overview Module # 4 - Wildlife and Nature Conservation Overview Component # 1 - Eastern Cape Wildlife and Nature Conservation Overview Module # 5 - Nelson Mandela Bay Metropole Component # 1 - Explore the Nelson Mandela Bay Metropole Module # 6 - Sarah Baartman District Municipality Component # 1 - Explore the Sarah Baartman District (Part 1) Component # 2 - Explore the Sarah Baartman District (Part 2) Component # 3 - Explore the Sarah Baartman District (Part 3) Component # 4 - Explore the Sarah Baartman District (Part 4) Module # 7 - Chris Hani District Municipality Component # 1 - Explore the Chris Hani District Module # 8 - Joe Gqabi District Municipality Component # 1 - Explore the Joe Gqabi District Module # 9 - Alfred Nzo District Municipality Component # 1 - Explore the Alfred Nzo District Module # 10 - OR Tambo District Municipality Component # 1 - Explore the OR Tambo District Eastern Cape Province Overview This course material is the copyrighted intellectual property of WildlifeCampus. -

A Phase 1 Archaeological Impact Assessment (Aia) for the Proposed Granor Passi Effluent Evaporation Ponds, Louterwater, Sarah Ba
A PHASE 1 ARCHAEOLOGICAL IMPACT ASSESSMENT (AIA) FOR THE PROPOSED GRANOR PASSI EFFLUENT EVAPORATION PONDS, LOUTERWATER, SARAH BAARTMAN DISTRICT MUNICIPALITY, KOUGA LOCAL MUNICIPALITY, EASTERN CAPE PROVINCE. Prepared for: SRK Consulting PO Box 21842 Port Elizabeth 6000 Tel: 041 509 4800 Fax: 041 509 4850 Contact person: Ms Karissa Nel Email: [email protected] Compiled by: Ms Celeste Booth t/a Booth Heritage Consulting 5 Queens Terrace 12 Chapel Street Grahamstown 6139 Tel: 082 062 4655 Email: [email protected] Contact person: Ms Celeste Booth Date: July 2016 1 CONTENTS 1. EXECUTIVE SUMMARY 3. 1.1. Purpose of the Study 3. 1.2. Brief Summary of Findings 3. 1.3. Recommendations 4. 1.4. Declaration of Independence and Qualifications 4. 2. BACKGROUND INFORMATION 5. 2.1. Applicant 5. 2.2. Consultant 5. 2.3. Terms of Reference 5. 3. HERITAGE LEGISLATIVE REQUIREMENTS 6. 4. BRIEF ARCHAEOLOGICAL BACKGROUND 8. 4.1. Early Stone Age (ESA) - 1.5 million to 250 000 years ago 9. 4.2. Middle Stone Age (MSA) – 250 000 – 30 000 years ago 10. 4.3. Later Stone Age (LSA) – 30 000 years ago – recent (100 years ago) 11. 4.4. Last 2 000 years – Khoekhoen Pastoralism 13. 4.5. Human Remains 14. 4.6. Rock Art (Paintings and Engravings) 14. 5. DESCRIPTION OF THE PROPERTY 15. 5.1. Location data 15. 5.2. Map 15. 6. ARCHAEOLOGICAL INVESTIGATION 21. 6.1. Methodology 21. 6.2. Results of the Archaeological Investigation 21. 7. DESCRIPTION OF SITES 29. 7.1. Middle Stone Age stone artefact occurrences 29. 8. COORDINATES AND SITES FOR THE PROPOSED SAND MINING ON ERF 220 AND ERF 252 OF THE FARM COEGAS RIVER MOUTH 303, COEGA, NELSON MANDELA METROPOLITAN MUNICIPALITY (NMMM), EASTERN CAPE PROVINCE. -

Thesis Sci 2020 Stander Johan.Pdf
University of Cape Town “Deriving a policy document towards an Early Warning System for Estuaries in South Africa: Case study Great Brak Estuary, Eden District, Southern Cape” Johan Stander Supervised by: Professor Isabelle Ansorge Head of Oceanography Department, University of Cape Town Associate Professor Juliet Hermes Manager, SAEON Egagasini node Associate Professor, Oceanography Department, University of Cape Town A thesis presented for the degree of Doctor of Philosophy in Oceanography University ofUniversity Cape Town of Cape Town June 2020 1 The copyright of this thesis vests in the author. No quotation from it or information derived from it is to be published without full acknowledgement of the source. The thesis is to be used for private study or non- commercial research purposes only. Published by the University of Cape Town (UCT) in terms of the non-exclusive license granted to UCT by the author. University of Cape Town Abstract South Africa’s estuaries and their surrounding communities are becoming increasingly vulnerable to storm surges and accompanied estuary flooding. These events are largely due to increasing severity of storm surges combined with growing housing and commercial developments. A particularly severe weather event in 2007/2008 highlighted the pressing need to understand the processes involved and the urgency to develop proactive response and management actions to mitigate the effects of future storm events on these coastal areas. Scientific research on estuarine flooding is limited not only for South Africa but within the international community as well and only recently has received committed attention from policy makers. It is clear that our current knowledge of South African estuary flooding events remains rudimentary; while necessary action to mitigate such events are poorly understood and planned. -

Great Brak Island – Measures for Flood Defence
GREAT BRAK ISLAND – MEASURES FOR FLOOD DEFENCE BY JT VILJOEN Thesis presented in fulfilment of the requirements for the degree of Master of Science in Engineering in the Faculty of Civil Engineering at Stellenbosch University Supervisor: Dr AK Theron Faculty of Engineering Department of Civil Engineering Division of Water and Environmental Engineering Stellenbosch University https://scholar.sun.ac.za Declaration By submitting this thesis electronically, I declare that the entirety of the work contained therein is my own, original work, that I am the sole author thereof (save to the extent explicitly otherwise stated), that reproduction and publication thereof by Stellenbosch University will not infringe any third party rights and that I have not previously in its entirety or in part submitted it for obtaining any qualification. Signature: …………………. Date: December 2017 Copyright © 2017 Stellenbosch University All rights reserved Page | i Stellenbosch University https://scholar.sun.ac.za Abstract This thesis firstly presents the analysis of large flood events, causing low-lying property in and around the Great Brak estuary to be inundated. The nature of several South African estuaries renders adjacent properties vulnerable to river and ocean related flooding. An important objective of the study is to develop a feasible form of flood defence for the Island, a residential development situated in the lower estuary basin, 250 m from the estuary mouth. The Island regularly experiences inundation due to high water levels in the estuary which have partially driven the need for the artificial manipulation of the estuary mouth berm using millions of litres of potable water. The lowest property on the Island is situated at +2.2 m MSL whereas the highest flood level ever recorded in the estuary was +2.9 m MSL. -

Environmental Scoping Report: Seafield Kleinemonde Eco-Estate DRAFT Coastal & Environmental Services
Coastal & Environmental Services THE PROPOSED ESTABLISHMENT OF AN ‘ECO- ESTATE’ DEVELOPMENT ADJACENT TO THE EAST KLEINEMONDE RIVER, EASTERN CAPE DRAFT ENVIRONMENTAL SCOPING REPORT Prepared by Coastal & Environmental Services P.O. Box 934 Grahamstown 6140 On behalf of Mr R Taylor Prepared for Approval by Department of Economic Affairs, Environment and Tourism Private Bag X5001 Greenacres Port Elizabeth 6057 25 October 2006 Environmental Scoping Report: Seafield Kleinemonde Eco-Estate DRAFT Coastal & Environmental Services TABLE OF CONTENTS 1 INTRODUCTION ........................................................................................................................................... 5 1.1 LIMITATIONS & ASSUMPTIONS........................................................................................................ 5 1.1.1 Limiting conditions ........................................................................................................................... 6 1.2 STUDY AREA, STUDY SITE AND STUDY TEAM .............................................................................. 6 1.3 GENERAL METHODOLOGY AND APPROACH ................................................................................ 6 2 THE ENVIRONMENTAL IMPACT ASSESSMENT (EIA) PROCESS ........................................................... 7 2.1 TERMS OF REFERENCE FOR THE ENVIRONMENTAL SCOPING STUDY .................................... 7 2.1.1 Role of the Environmental Consultant or Environmental assessment Practitioner .......................... 8 2.1.2 -
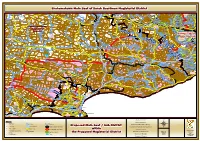
EC Sbmain 032018 Grahamst
!C !.C^ !C$ñ ^!C ^ ^ !C !C !C !C !C ^ !C !C ^ !C^ !C !C !C !C !C^ !C ñ !C !C !C !C !C !C ^ !C !C !C ^$ !C ^ !C !C !C !C !C !C ^ ^!C !C !C !C ñ !C !C !C !C !C !C !C !C !C !. !C ^ ^ !C ñ !C !C !C $ !C !C !C!C ^ !C !C !C ^ !C !C ñ !C !C !C ñ!.C^ !C !C ñ !C ^ !C !C ^ ^ ^ !C !C !C !C !C !C !C !C !C ^ !C !C !C ñ !C !C ^ !C ñ !C !C !C ñ !C !C !C !C !C !C !C !C !C !C ñ !C !C ^ !C ^ !C !C !C ñ!C ^!C !. ^ !C !C !C ^ !C !C ^ ñ $ ^$!C ^ !C !C !C !C !C !C !C !C !C !C !C !C !. !C !.^ $ !C !C ñ !C !C ^ !C !C !C $ ^ !C !C $!C !C !C ñ !. $ !C !C !C !C!C !C ñ!C. ^ ^ ^ !C $!.!C^ !C !C !C !C !C !C !C !C !C !C !C !C !C!C !. !C !C !C !C !.^!C !C !C !C !C ñ !C !Cñ^ !C !C ñ !.^ !C !C !C !C !C !C ^!C!C ^$ !C ^ñ!C ñ !C ñ!C!.^ !C !. ^ !C !C ^ñ !. !C $^ ñ^!C ^ ^ !C ^ ñ ^ !C !C !C !C !C !C !C !C !C ^ !C !C !C !C !C !C !. !C ^ !C $ !. ñ!C !C !C ^ !C.^ ñ !C !C !C !C !C !C !C !C ^!.$C !. !. !C ^ ^ !C !C^ !C !C!. !C !C !C ñ!C !C !. $!C^ !C !C !C !C !C !.