Use of Endophytic Bacteria Naturally Associated with Cestrum Nocturnum for Fusarium Wilt Biocontrol and Enhancement of Tomato Growth
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Weedsoc.Org.Au
THE WEED SOCIETY OF NEW SOUTH WALES Inc. Website: www.nswweedsoc.org.au Seminar Papers WEEDS – WOE to GO IV Wednesday 6 September 2006 Metcalfe Auditorium State Library of NSW Macquarie Street , SYDNEY Sponsors Collated / Edited by Copies of this publication are available from: Dr Stephen Johnson THE WEED SOCIETY & Bob Trounce OF NEW SOUTH WALES Inc. PO Box 438 WAHROONGA NSW 2076 THE WEED SOCIETY OF NEW SOUTH WALES Inc. ACKNOWLEDGMENTS Seminar Organising Committee Lawrie Greenup (chair) Mike Barrett Bertie Hennecke Luc Streit Coordinator power point presentations Erica McKay Welcome to speakers and delegates Warwick Felton (President) Summary of the day’s presentations Mike Barrett Collation and preparation of proceedings Stephen Johnson Bob Trounce The committee thanks all who took part and attended the seminar and particularly the speakers for their presentations and supply of written documents for these proceedings. THE WEED SOCIETY OF NEW SOUTH WALES Inc. SEMINAR SERIES: WEEDS WOE TO GO IV “Poisonous and Allergenic Plants Where are they?” Date: Wednesday 6th September 2006 Location: The Metcalfe Auditorium The State Library of NSW Macquarie Street Sydney Time Topic Speaker 9.00 – 9.30 am REGISTRATION & MORNING TEA 9.30 – 9.40 am Welcome Warwick Felton 9.40 – 10.30 am Weeds that make you sick Rachel McFadyen 10.30 – 11.20 am Poisonous, prickly, parasitic, pushy? John Virtue Prioritising weeds for coordinated control programs” 11.20 – 1130 am break 11.30 – 11.50 am Parietaria or Asthma Weed Sue Stevens Education & incentive project -

Appendix Color Plates of Solanales Species
Appendix Color Plates of Solanales Species The first half of the color plates (Plates 1–8) shows a selection of phytochemically prominent solanaceous species, the second half (Plates 9–16) a selection of convol- vulaceous counterparts. The scientific name of the species in bold (for authorities see text and tables) may be followed (in brackets) by a frequently used though invalid synonym and/or a common name if existent. The next information refers to the habitus, origin/natural distribution, and – if applicable – cultivation. If more than one photograph is shown for a certain species there will be explanations for each of them. Finally, section numbers of the phytochemical Chapters 3–8 are given, where the respective species are discussed. The individually combined occurrence of sec- ondary metabolites from different structural classes characterizes every species. However, it has to be remembered that a small number of citations does not neces- sarily indicate a poorer secondary metabolism in a respective species compared with others; this may just be due to less studies being carried out. Solanaceae Plate 1a Anthocercis littorea (yellow tailflower): erect or rarely sprawling shrub (to 3 m); W- and SW-Australia; Sects. 3.1 / 3.4 Plate 1b, c Atropa belladonna (deadly nightshade): erect herbaceous perennial plant (to 1.5 m); Europe to central Asia (naturalized: N-USA; cultivated as a medicinal plant); b fruiting twig; c flowers, unripe (green) and ripe (black) berries; Sects. 3.1 / 3.3.2 / 3.4 / 3.5 / 6.5.2 / 7.5.1 / 7.7.2 / 7.7.4.3 Plate 1d Brugmansia versicolor (angel’s trumpet): shrub or small tree (to 5 m); tropical parts of Ecuador west of the Andes (cultivated as an ornamental in tropical and subtropical regions); Sect. -

Green Cestrum
OCTOBER 2008 PRIMEFACT 718 REPLACES AGFACT P7.6.44 Green cestrum Neil Griffiths Impact District Agronomist, NSW DPI, Tocal, Paterson Green cestrum is a vigorous plant that can out- compete other vegetation. Green cestrum is toxic to Dr Chris Bourke animals including cattle, sheep, horse, pigs, poultry Principal Research Scientist (Poisonous Plants), and humans. NSW DPI, Orange Agricultural Institute Habitat Green cestrum is normally found along watercourses and in non-crop areas where it usually grows in small to medium-sized thickets. Introduction Distribution Green cestrum (Cestrum parqui) is a large poisonous shrub belonging to the Solanaceae family. In NSW, green cestrum is found in the Hunter Valley, The plant is also known as green poison berry or the outer metropolitan areas of Sydney, the North Coast and the north-west, central west and south- Chilean cestrum. west of the State. Green cestrum was originally introduced into Australia from South America as an ornamental shrub for Description gardens. Since that time, it has become naturalised in Green cestrum is a medium-sized perennial areas of south-eastern Queensland, eastern New South shrub growing 2–3 m (Figure 2). It usually has many Wales (NSW) and parts of Victoria and South Australia. light-green, brittle stems. Figure 1. Green cestrum is a Class 3 noxious weed in NSW. Photo: G. Wisemantel. Leaves Fruit The shiny-green leaves are 20–30 mm wide and Clusters of shiny, black, egg-shaped berries 80–100 mm long. They have smooth edges, are 7–10 mm long are produced during summer and pointed at each end and are arranged alternately autumn (Figure 4). -

Antibacterial and Antifungal Activities of Cestrum Parqui Saponins: Possible Interaction with Membrane Sterols
International Research Journal of Plant Science (ISSN: 2141-5447) Vol. 3(1) pp. 001-007, January, 2012 Available online http://www.interesjournals.org/IRJPS Copyright © 2012 International Research Journals Full length Research Paper Antibacterial and antifungal activities of Cestrum parqui saponins: possible interaction with membrane sterols Dorsaf Ben Ahmed 1, Ikbal Chaieb 4, Karima Belhadj Salah 2, Habib Boukamcha 3, Hichem Ben Jannet 3, Zine Mighri 3, and Mejda Daami-Remadi 4* 1Institut Supérieur Agronomique de Chott-Mariem, 4042 Chott-Mariem, Université de Sousse, Tunisia 2Laboratoire des Maladies Transmissibles et des Substances Biologiquement Actives, Faculté de Pharmacie de Monastir, 5000, Université de Monastir, Tunisia 3Laboratoire des Substances Naturelles et de Synthèse Organique, Faculté des Sciences de Monastir, 5000, Université de Monastir, Tunisia 4Centre Régional des Recherches en Horticulture et Agriculture Biologique, 4042 Chott-Mariem, Université de Sousse, Tunisia Accepted 10 November, 2011 Cestrum parqui L´Hér. (Solanaceae) is used as ornamental plant in Tunisia. This plant is rich in saponin content which was largely described as a fungicidal compound synthesized by plants for defence purposes. The aim of the present work is to assess the activity of the crude saponic extract (CSE) on several bacterial and fungal agents and to study the interaction between saponin and membrane sterols in relation with their eventual inhibitory activities. Two Gram-positive (Pseudomonas aeruginosa and Escherichia coli) and two Gram-negative ( Staphylococcus aureus and Enterococcus faecalis ) bacteria were tested. No bacterial species was found to be sensitive to C. parqui saponins even with the highest CSE concentration used (100 mg/ml). The antifungal activity was confirmed against two plant pathogens ( Fusarium solani and Botrytis cinerea ) and one antagonistic agent (Trichoderma viride ). -

2641-3182 08 Catalogo1 Dicotyledoneae4 Pag2641 ONAG
2962 - Simaroubaceae Dicotyledoneae Quassia glabra (Engl.) Noot. = Simaba glabra Engl. SIPARUNACEAE Referencias: Pirani, J. R., 1987. Autores: Hausner, G. & Renner, S. S. Quassia praecox (Hassl.) Noot. = Simaba praecox Hassl. Referencias: Pirani, J. R., 1987. 1 género, 1 especie. Quassia trichilioides (A. St.-Hil.) D. Dietr. = Simaba trichilioides A. St.-Hil. Siparuna Aubl. Referencias: Pirani, J. R., 1987. Número de especies: 1 Siparuna guianensis Aubl. Simaba Aubl. Referencias: Renner, S. S. & Hausner, G., 2005. Número de especies: 3, 1 endémica Arbusto o arbolito. Nativa. 0–600 m. Países: PRY(AMA). Simaba glabra Engl. Ejemplares de referencia: PRY[Hassler, E. 11960 (F, G, GH, Sin.: Quassia glabra (Engl.) Noot., Simaba glabra Engl. K, NY)]. subsp. trijuga Hassl., Simaba glabra Engl. var. emarginata Hassl., Simaba glabra Engl. var. inaequilatera Hassl. Referencias: Basualdo, I. Z. & Soria Rey, N., 2002; Fernández Casas, F. J., 1988; Pirani, J. R., 1987, 2002c; SOLANACEAE Sleumer, H. O., 1953b. Arbusto o árbol. Nativa. 0–500 m. Coordinador: Barboza, G. E. Países: ARG(MIS); PRY(AMA, CAA, CON). Autores: Stehmann, J. R. & Semir, J. (Calibrachoa y Ejemplares de referencia: ARG[Molfino, J. F. s.n. (BA)]; Petunia), Matesevach, M., Barboza, G. E., Spooner, PRY[Hassler, E. 10569 (G, LIL, P)]. D. M., Clausen, A. M. & Peralta, I. E. (Solanum sect. Petota), Barboza, G. E., Matesevach, M. & Simaba glabra Engl. var. emarginata Hassl. = Simaba Mentz, L. A. glabra Engl. Referencias: Pirani, J. R., 1987. 41 géneros, 500 especies, 250 especies endémicas, 7 Simaba glabra Engl. var. inaequilatera Hassl. = Simaba especies introducidas. glabra Engl. Referencias: Pirani, J. R., 1987. Acnistus Schott Número de especies: 1 Simaba glabra Engl. -

Garden Plants Poisonous to People
N NO V E M B E R 2 0 0 6 P R I M E F A C T 3 5 9 ( R E P L A C E S A G F A C T P 7 . 1 . 1 P O I S O N O U S P L A N T S I N T H E G A R D E N) Garden plants poisonous to people Annie Johnson Table 1. Toxicity rating for Tables 2−7. Weeds Project Officer Rating Toxicity Stephen Johnson Mildly toxic. Mild symptoms may occur if large * Weed Ecologist quantities are eaten. Toxic. Causes discomfort and irritation but not Weeds Unit, Biosecurity Compliance and Mine ** Safety, Orange dangerous to life. Highly toxic. Capable of causing serious illness *** or death. Introduction There are a range of garden plants that are considered poisonous. Poisonings and deaths from garden plants Poisoning are rare as most poisonous plants taste unpleasant Poisoning from plants may occur from ingesting, and are seldom swallowed (see toxicity). However, it is inhalation or direct contact. best to know which plants are potentially toxic. Symptoms from ingestion include gastroenteritis, It is important to remember that small children are diarrhoea, vomiting, nervous symptoms and in serious often at risk from coloured berries, petals and leaves cases, respiratory and cardiac distress. Poisoning that look succulent. This does not mean that all these by inhalation of pollen, dust or fumes from burning poisonous plants should be avoided or removed from plants can cause symptoms similar to hay fever or the garden. It is best to teach children never to eat asthma. -

The First Report of Root-Knot Nematode on Cestrum Nocturnum in Ninawa, Iraq
Plant Archives Volume 20 No. 2, 2020 pp. 6778-6780 e-ISSN:2581-6063 (online), ISSN:0972-5210 THE FIRST REPORT OF ROOT-KNOT NEMATODE ON CESTRUM NOCTURNUM IN NINAWA, IRAQ Firas K. Aljuboori1* and Asmaa Mansour Al-Hakeem2 1*Department of Plant Protection, College of Agriculture and Forestry, University of Mosul, Iraq. 2Department of Microbiology, College of Science, Al-Karkh University of Science, Iraq Abstract Night-blooming jasmine Cestrum nocturnum is one of the common ornamental plants in Iraq. It is widely planted in the house gardens, public gardens and nurseries. A survey was carried out during 2019 to assist the host range of Meloidogyne spp. in ornamental plants. Samples were collected from house gardens, public gardens and nurseries in Ninawa governorate - Iraq. The root-knot nematode was identified by the root symptoms and the presence of females and egg mass. Nematode species was identified depending on the adult female perineal pattern. The most common symptoms of the infected plants were stunting, foliage yellowing, lack of vigor and wilt during the hot summer. Nematode infection was indexed depending on the root galling using a 0-5 scale. Root-knot density and severity were estimated according to both natural and artificial infection. House and public gardens have a higher infection percentage than nurseries (10.4, 1.9) while the severity is higher in nurseries (0.142, 0.35). According to the best of the researchers’ knowledge, this is the first report of Meloidogyne javanica on night-blooming jasmine in Iraq. Key words: root-knot nematode, Cestrum nocturnum, Meloidogyne javanica. Introduction ornamental plants and herbs (Ralmi et al., 2016). -

Delicate, Fragrant, Lady of the Night- a Medicinal Received: 04-09-2016 Accepted: 10-10-2016 Gift
Journal of Medicinal Plants Studies 2016; 4(6): 13-17 ISSN 2320-3862 JMPS 2016; 4(6): 13-17 © 2016 JMPS Delicate, fragrant, lady of the night- A medicinal Received: 04-09-2016 Accepted: 10-10-2016 gift Amin Shaista Department of Pharmaceutical Amin Shaista and Parle Amrita Chemistry, Delhi Pharmaceutical Science and Research University, Abstract New Delhi India Night blooming jasmine, botanically known as Cestrum nocturnum is an evergreen shrub that grows in Parle Amrita tropical and sub-tropical regions throughout the world. Cestrum nocturnum is a popular ornamental plant Department of Pharmaceutical due to its showy and fragrant white flowers. It is also used as a hedge plant and cultivated as a medicinal Chemistry, Delhi Pharmaceutical plant. The medicinal properties of night blooming jasmine include antioxidant, anti-hyperlipidemic, Science and Research University, hepatoprotective, analgesic, antibacterial, antifungal, anti-convulsant, anti-HIV and larvicidal activities. New Delhi India The present paper reviews the geographical distribution, history, cultivation, uses, side effects, synonyms, botanical description, taxonomical classification, phytochemical constituents and pharmacological activities. Keywords: Cestrum nocturnum, antibacterial, antioxidant, anti-inflammatory, larvicidal Introduction Cestrum nocturnum is a garden shrub from the family Solanaceae, commonly known as "lady of the night" which is used as a remedy for different health disorders. This sprawling shrub has glossy simple leaves, vine like stems, greenish-creamy white tubular flowers and fleshy berries. The berries are marfil white or aubergine in colour. The species name ‘nocturnum’ refers to the species’ habit of opening its small, heavily-scented flowers at night. The flowers release powerful sweet perfume at night. It is made into a rare attar (raat ki rani) which is used in Indian and Middle East perfumery. -
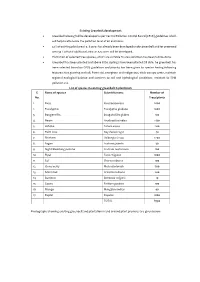
CPCB) Guidelines Which Will Help to Attenuate the Pollution Level of Air and Noise
Existing Greenbelt development Greenbelt is being/will be developed as per Central Pollution Control Board (CPCB) guidelines which will help to attenuate the pollution level of air and noise. 33 % of existing plant area i.e. 8 acres has already been developed under greenbelt and for proposed area 33 % of total additional area i.e. 5.5 acres will be developed. Plantation of selected tree species, which are suitable to area condition has been/will be done. Greenbelt has been planted and above 8750 saplings have been planted till date. he greenbelt has been selected based on CPCB guidelines and priority has been given to species having following features; fast growing and tall, Perennial, evergreen and indigenous, thick canopy cover, maintain regional ecological balance and conform to soil and hydrological conditions, resistant to SPM pollution etc. List of species in existing greenbelt & plantation S. Name of species Scientific name Number of No. Trees/plants 1. Ficus Ficus benjamina 2000 2. Eucalyptus Eucalyptus globulus 1000 3. Bougenvillia Bougainvillea glabra 100 4. Neem Azadirachta indica 1130 5. Ashoka Saraca asoca 200 6. Palm Tree Roystonea regia 40 7. Shisham Dalbergia sissoo 1200 8. Sagon Tectona grandis 30 9. Night blooming jasmine Cestrum nocturnum 100 10. Pipal Ficus religiosa 1000 11. Sal Shorea robusta 100 12. China berry Melia azedarach 500 13. Silver Oak Grevillea robusta 200 14. Bamboo Bambusa vulgaris 10 15. Guava Psidium guajava 100 16. Mango Mangifera indica 40 17. Poplar Populus 1000 TOTAL 8750 Photographs showing existing greenbelt and plantation in and around plant premises are given below: Action plan for greenbelt development for proposed area The rate of pollutant removal is found to increase linearly as the concentration of the pollutant increases over the range of concentration that are encountered in ambient air and which are low enough not to cause stomatal closure. -

Cestrum Nocturnum Global Invasive
FULL ACCOUNT FOR: Cestrum nocturnum Cestrum nocturnum System: Terrestrial Kingdom Phylum Class Order Family Plantae Magnoliophyta Magnoliopsida Solanales Solanaceae Common name onaona Iapana (Hawaiian), ike he po (Niuean), thauthau (Fijian), ali'I o le po (Samoan), teine 'o le po (Samoan), galan de noche (Spanish), ye xiang shu (Mandarin), laukau po'uli (Tongan), fafine o te po (Tuvaluan), ala aumoe (Hawaiian), kupaoa (Hawaiian), thauthau ni mbongi (Fijian), tiare ariki va'ine (Cook Islands), jonoul ruo awa (Marshallese), ai pua e pogi (Rotuman), ariki-va'ine (Cook Islands), night jessamine (English), kara (Fijian), dama de noche (Chamorro), night queen (English), lady of the night (English), night cestrum (English), night-blooming jasmine (English), night-flowering cestrum (English), dama di noche (Chamorro), night-flowering jasmine (English), queen of the night (English) Synonym Cestrum parqui Similar species Cestrum parqui Summary Cestrum nocturnum commonly known as queen of the night, is a popular ornamental species widely distributed for its strongly fragrant flowers that bloom at night. Having bird-dispersed seeds and the ability to reproduce vegetatively has resulted in escapes from cultivation, where in some areas it aggressively colonises disturbed sites such as road edges and forest gaps forming dense impenetrable thickets and resulting in competition with and displacement of native plant species. C. nocturnum is also known to be poisonous if ingested, forming a risk to grazing livestock and has been known to produce hay-fever like symptoms in some people. view this species on IUCN Red List Global Invasive Species Database (GISD) 2021. Species profile Cestrum nocturnum. Pag. 1 Available from: http://www.iucngisd.org/gisd/species.php?sc=851 [Accessed 07 October 2021] FULL ACCOUNT FOR: Cestrum nocturnum Species Description Cestrum nocturnum is a glabrous shrub that grows from 1 to 5 m tall (depending on location) with ovate-oblong, petiolate, and obtuse leaves mostly 7 - 20 cm long (Webb et al., 1988; Tharman et al., 1994; Zhang et al., 1994). -

47 in Vitro Nematicidal Activity of Aqueous Plant Extracts
IN VITRO NEMATICIDAL ACTIVITY OF AQUEOUS PLANT EXTRACTS ON CHILEAN POPULATIONS OF XIPHINEMA AMERICANUM SENSU LATO V. Insunza,1 E. Aballay, 2 and J. Macaya2 Swedish University of Agricultural Sciences, Department of Ecology and Crop Production Science, P.O. Box 7043, SE-750 07 Uppsala, Sweden1 and Universidad de Chile, Facultad de Ciencias Agronómicas, Departamento de Sanidad Vegetal, Casilla 1004, Santiago, Chile. 2 ABSTRACT Insunza, V., E. Aballay, and J. Macaya. 2001. In vitro nematicidal activity of aqueous plant extracts on Chilean populations of Xiphinema americanum sensu lato. Nematropica 31:47-54. In a search for alternatives to chemicals in nematode management, the nematicidal activity of 30 plant species (9 native and 21 naturalized in Chile) was tested on a Chilean population of Xiphinema americanum sensu lato, an economically important pathogen in grapevine. Aqueous extracts were test- ed in a standard concentration (S) at 1:4 W/V of fresh plant material, or 1:20 W/V of air-dried plant parts; and 21 plants, were also tested at 25% of S. Distilled water was used as the control. Nematicidal activity was evaluated by nematode immobility after 24-48 h of immersion in the extracts, followed by 24 h in distilled water. At the standard concentration, all the extracts showed nematicidal or nemato- static activity after 24 h of exposure, but this effect decreased in the dilute treatment of 33% of the extracts, mostly top extracts. Nematode immobility was observed at 25% of S, after 24 h, with plant extracts in 18 out of 21 plant species including: Tagetes erecta, T. -

Cestrum Parqui Global Invasive Species Database (GISD)
FULL ACCOUNT FOR: Cestrum parqui Cestrum parqui System: Terrestrial Kingdom Phylum Class Order Family Plantae Magnoliophyta Magnoliopsida Solanales Solanaceae Common name palqui (English), duraznillo negro (English), willow-leafed jessamine (English), duraznillo (English), duraznillo hediondo (English), green cestrum (English), mala yerba (English), palque (English), hediondilla (English), coerana (Portuguese), coeirana (English), quina del campo (Spanish), Chilean flowering jassamine (English), green poison berry (English), Chilean cestrum (English), willow jasmine (English) Synonym Similar species Cestrum nocturnum, Cestrum elegans, Cestrum aurantiacum Summary Cestrum parqui is a shrub of the Solanaceae family that is a toxic plant and has become an invasive weed. It prefers moist habitats but is commonly found along roadsides, and neglected, disturbed, and abandoned sites. This species exhibits vigorous growth and out competes most native vegetation. With its extensive, shallow root system C. parqui is extremely difficult to eradicate, however there are various control methods available. C. parqui is toxic and is responsible for the death livestock around the world annually. It is just as dangerous to humans. view this species on IUCN Red List Global Invasive Species Database (GISD) 2021. Species profile Cestrum parqui. Pag. 1 Available from: http://www.iucngisd.org/gisd/species.php?sc=850 [Accessed 06 October 2021] FULL ACCOUNT FOR: Cestrum parqui Species Description QDNRM (2005) states that, \"Cestrum parqui is an erect, perennial shrub to 3m high, with one or more stems emerging from each crown. The young stems are whitish; older stems are darker, striated at the base and mottled above. The leaves are alternate, up to 12cm long and 2.5cm wide, and have an unpleasant odor when crushed.