Vocal Mimicry in Spotted Bowerbirds Is Associated with an Alarming Context
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Does Gray Squirrel (Sciurus Carolinensis) Response to Heterospecific Alarm Calls Depend on Familiarity Or Acoustic Similarity?
Ethology RESEARCH PAPER Does Gray Squirrel (Sciurus carolinensis) Response to Heterospecific Alarm Calls Depend on Familiarity or Acoustic Similarity? Cynthia M. Getschow, Pearl Rivers, Samantha Sterman, David C. Lumpkin & Keith A. Tarvin Department of Biology, Oberlin College, Oberlin, OH, USA Correspondence Abstract Keith A. Tarvin, Department of Biology, Oberlin College, 119 Woodland Street, In habitats in which multiple species are prey to the same predators, indi- Oberlin, OH 44074, USA. viduals can greatly benefit from recognizing information regarding preda- E-mail: [email protected] tors that is provided by other species. Past studies have demonstrated that various mammals respond to familiar heterospecific alarm calls, but Received: January 28, 2013 whether acoustic similarity to a familiar call can prompt a mammal’s Initial acceptance: May 14, 2013 recognition of an unfamiliar call has yet to be shown. We presented alarm Final acceptance: August 2, 2013 calls to free-ranging eastern gray squirrels (Sciurus carolinensis) and (D. Zeh) recorded behavioral changes in vigilance and antipredatory response. Playbacks included alarm calls of a sympatric bird (American robin, Turdus doi: 10.1111/eth.12145 migratorius), an allopatric bird with a call structure similar to that of the robin (common blackbird, Turdus merula), and an allopatric bird with a distinct call structure (New Holland honeyeater, Phylidonyris novaehollan- diae). Squirrels responded significantly more frequently to squirrel alarm calls (positive control) than to robin song (negative control) or honeyeater calls. Squirrel response to robin and blackbird alarm calls was statistically similar to their response to squirrel alarm calls, indicating that squirrels responded to those alarm calls as if they provided information about the presence of predators. -

Aspirating Smoke Detection APPLICATIONS GUIDE: ASPIRATING SMOKE DETECTION Aspirating Smoke Detection
APPLICATIONS GUIDE Aspirating Smoke Detection APPLICATIONS GUIDE: ASPIRATING SMOKE DETECTION Aspirating Smoke Detection Contents Aspirating Smoke Detection ...............................................................................3 Design Best Practices .......................................................................................21 Codes and Standards .......................................................................................4 More on Hot Aisle/ Cold Aisle Configurations .................................................22 Definitions ............................................................................................................4 Coordination and Interface with other Systems ..............................................26 United States Definitions and Requirements .....................................................4 Common Issues / Application Troubleshooting .............................................26 Requirements of SFD systems according to NFPA 72 .....................................4 Telecommunications .......................................................................................28 Requirements of EWFD systems according to NFPA 76 .................................4 Application Overview.........................................................................................28 Requirements of VEWFD systems according to NFPA 76 ...............................5 Benefits of Aspirating Smoke Detection ..........................................................29 European EN 54-20 Requirements -

Birds in and Around Blackall
Birds in and around Blackall Apostle bird Eastern Rosella Red backed Kingfisher Australasian Grebe Emu Red-backed Fairy-wren Australasian Shoveler Eurasian Coot Red-breasted Button Quail Australian Bustard Forest Kingfisher Red-browed Pardalote Australian Darter Friary Martin Red-capped Robin Australian Hobby Galah Red-chested Buttonquail Australian Magpie Glossy Ibis Red-tailed Black-Cockatoo Australian Magpie-lark Golden-headed Cisticola Red-winged Parrot Australian Owlet-nightjar Great (Black) Cormorant Restless Flycatcher Australian Pelican Great Egret Richard’s Pipit Australian Pipit Grey (White) Goshawk Royal Spoonbill Australian Pratincole Grey Butcherbird Rufous Fantail Australian Raven Grey Fantail Rufous Songlark Australian Reed Warbler Grey Shrike-thrush Rufous Whistler Australian White Ibis Grey Teal Sacred Ibis Australian Ringneck (mallee form) Grey-crowned Babbler Sacred Kingfisher Australian Wood Duck Grey-fronted Honeyeater Singing Bushlark Baillon’s Crake Grey-headed Honeyeater Singing Honeyeater Banded Honeyeater Hardhead Southern Boobook Owl Barking Owl Hoary-headed Grebe Spinifex Pigeon Barn Owl Hooded Robin Spiny-cheeked Honeyeater Bar-shouldered Dove Horsfield's Bronze-Cuckoo Splendid Fairy-wren Black Falcon House Sparrow Spotted Bowerbird Black Honeyeater Inland Thornbill Spotted Nightjar Black Kite Intermediate Egret Square-tailed kite Black-chinned Honeyeater Jacky Winter Squatter Pigeon Black-faced Cuckoo-shrike Laughing Kookaburra Straw-necked Ibis Black-faced Woodswallow Little Black Cormorant Striated Pardalote -

Wednesday, March 14
Antipredator behavior Antipredator behavior Nonsocial species may rely on crypsis, aposematism or mimicry scorpionfish lonomia moth katydid Antipredator behavior Testing adaptations Nonsocial species may rely on We presume these are crypsis, aposematism or mimicry adaptations to predation, but how to be sure? against conspicuous background Catocala spp. Detection index Detection against cryptic background Müllerian mimicry: unpalatable species converge Batesian mimicry: palatable Trials mimic unpalatable model Testing adaptations What about the aposematic hindwings of Catocala spp.? Blue jays trained on: Then presented with: Result: Cryptic hindwings Aposematic hindwings Startled! Aposematic hindwings Aposematic hindwings Not startled Aposematic hindwings Different aposematic hindwings Startled! How does this affect hindwing coloration across the genus? 1 Scarlet kingsnake, Lampropeltis triangulum Eastern coral snake, Micrurus fulvius Scarlet kingsnake, Lampropeltis triangulum Eastern coral snake, Micrurus fulvius Non-venomous Highly venomous Hypothesis Bright coloration with striking Bright coloration with striking Kingsnakes are protected from predators banding patterns banding patterns because they mimic venomous coral snakes (red + black) (red + yellow) Kingsnakes ONLY Kingsnakes Coral snakes Coral snakes and Hypothesis Kingsnakes Kingsnakes are protected from predators because they mimic venomous coral snakes Scarlet kingsnake, Lampropeltis triangulum Eastern coral snake, Micrurus fulvius Scarlet kingsnake, Lampropeltis triangulum -

A LIST of the VERTEBRATES of SOUTH AUSTRALIA
A LIST of the VERTEBRATES of SOUTH AUSTRALIA updates. for Edition 4th Editors See A.C. Robinson K.D. Casperson Biological Survey and Research Heritage and Biodiversity Division Department for Environment and Heritage, South Australia M.N. Hutchinson South Australian Museum Department of Transport, Urban Planning and the Arts, South Australia 2000 i EDITORS A.C. Robinson & K.D. Casperson, Biological Survey and Research, Biological Survey and Research, Heritage and Biodiversity Division, Department for Environment and Heritage. G.P.O. Box 1047, Adelaide, SA, 5001 M.N. Hutchinson, Curator of Reptiles and Amphibians South Australian Museum, Department of Transport, Urban Planning and the Arts. GPO Box 234, Adelaide, SA 5001updates. for CARTOGRAPHY AND DESIGN Biological Survey & Research, Heritage and Biodiversity Division, Department for Environment and Heritage Edition Department for Environment and Heritage 2000 4thISBN 0 7308 5890 1 First Edition (edited by H.J. Aslin) published 1985 Second Edition (edited by C.H.S. Watts) published 1990 Third Edition (edited bySee A.C. Robinson, M.N. Hutchinson, and K.D. Casperson) published 2000 Cover Photograph: Clockwise:- Western Pygmy Possum, Cercartetus concinnus (Photo A. Robinson), Smooth Knob-tailed Gecko, Nephrurus levis (Photo A. Robinson), Painted Frog, Neobatrachus pictus (Photo A. Robinson), Desert Goby, Chlamydogobius eremius (Photo N. Armstrong),Osprey, Pandion haliaetus (Photo A. Robinson) ii _______________________________________________________________________________________ CONTENTS -

Birds on Farms
Border Rivers Birds on Farms INTRODUCTION Grain & Graze Forty-seven mixed farms within nine regions around Australia took part in BR All collecting ecological data for the biodiversityBorder section of the nationalRivers Grain Region and Graze project. Table 1 Bird Survey Statistics Farms Farms Native bird species 105 183 This factsheet outlines the results from bird surveys, conducted in Spring 2006 and Autumn 2006-2007, on five farms within the Border Rivers Region in Introduced bird species 1 6 Queensland and New South Wales. Listed Threatened Species** 7 33 NSW Listed Species 1 11 Four paddock types were surveyed, Crop, Rotation, Pasture and Remnant (see overleaf for link to methods). Although, more bird species were recorded Priority Species 3 15 in remnant vegetation, birds were also frequently observed in other land use ** State and/or Federally types (Table 2). Table 2 List of bird species recorded in each paddock in all of the 5 Border Rivers Region farms. Food Food Common Name Preference Crop Rotation Pasture RemnantCommon Name Preference Crop Rotation Pasture Remnant Grey-crowned Babbler (N) I 1 3 1 2 3 4 Brown Goshawk IC 4 Varied Sittella I 3 White-faced Heron IC 3 White-winged Triller IGF 3 3 4 5 White-necked Heron IC 3 Striped Honeyeater IN 1 3 1 3 2 3 4 Olive-backed Oriole IF 3 Blue-faced Honeyeater IN 1 1 2 3 5 Pacific Black Duck IG 3 5 Brown Honeyeater IN 5 Little Button-quail IG 3 Red-winged Parrot G 1 1 2 5 Noisy Miner IN 4 1 2 3 4 5 3 1 2 3 4 5 Wedge-tailed Eagle C 4 2 3 5 Little Friarbird IN 1 3 Brown Thornbill I -

Multimodal Alarm Behavior in Urban and Rural Gray Squir-Rels Studied By
Current Zoology 56 (3): 313−326, 2010 Multimodal alarm behavior in urban and rural gray squir- rels studied by means of observation and a mechanical robot Sarah R. PARTAN*, Andrew G. FULMER, Maya A. M. GOUNARD, Jake E. REDMOND Hampshire College School of Cognitive Science, Amherst, MA 01002 Massachusetts, USA Abstract Urbanization of animal habitats has the potential to affect the natural communication systems of any species able to survive in the changed environment. Urban animals such as squirrels use multiple signal channels to communicate, but it is un- known how urbanization has affected these behaviors. Multimodal communication, involving more than one sensory modality, can be studied by use of biomimetic mechanical animal models that are designed to simulate the multimodal signals and be pre- sented to animal subjects in the field. In this way the responses to the various signal components can be compared and contrasted to determine whether the multimodal signal is made up of redundant or nonredundant components. In this study, we presented wild gray squirrels in relatively urban and relatively rural habitats in Western Massachusetts with a biomimetic squirrel model that produced tail flags and alarm barks in a variety of combinations. We found that the squirrels responded to each unimodal component on its own, the bark and tail flag, but they responded most to the complete multimodal signal, containing both the acoustic and the moving visual components, providing evidence that in this context the signal components are redundant and that their combination elicits multimodal enhancement. We expanded on the results of Partan et al. (2009) by providing data on sig- naling behavior in the presence and absence of conspecifics, suggesting that alarm signaling is more likely if conspecifics are present. -

An Inverse Relationship Between Decoration and Food Colour Preferences in Satin Bowerbirds Does Not Support the Sensory Drive Hypothesis
ANIMAL BEHAVIOUR, 2006, 72, 1125e1133 doi:10.1016/j.anbehav.2006.03.015 An inverse relationship between decoration and food colour preferences in satin bowerbirds does not support the sensory drive hypothesis GERALD BORGIA*† & JASON KEAGY† *Department of Biology, University of Maryland, College Park yDepartment of Behavior, Ecology, Evolution and Systematics Program, University of Maryland, College Park (Received 20 September 2005; initial acceptance 25 October 2005; final acceptance 30 March 2006; published online 26 September 2006; MS. number: A10246) Male bowerbirds collect and decorate their bowers with coloured objects that influence female choice. A recent version of the sensory drive hypothesis claims that female food colour preferences have driven the evolution of female preferences for the colour of male display traits. This hypothesis predicts a positive correlation between male display and food colour preferences. A positive correlation between food and decoration preferences could also arise because of sensory biases built into bowerbirds or the environment. Here we test hypotheses that (1) male and female satin bowerbirds show well-defined food colour prefer- ences, (2) these preferences correlate with independently assessed preferences for bower decorations, and, in a cross-species comparison, (3) food items were used as the first bower decorations. We found that male and female satin bowerbirds, Ptilonorhynchus violaceus, preferentially use long wavelength and were colours as food items. Male decoration preferences were biased towards colours of short wavelength and were negatively correlated with food colour preferences. Our reconstruction of ancestral character states is most consistent with the hypothesis that the original bower decorations were inedible objects and were thus unlikely to have been dual-use traits that also functioned as food items. -

Redalyc.Egg-Guarding Behavior of the Treehopper Ennya Chrysura
Revista de Biología Tropical ISSN: 0034-7744 [email protected] Universidad de Costa Rica Costa Rica Miranda, Ximena Egg-guarding behavior of the treehopper Ennya chrysura (Hemiptera: Membracidae): female aggregations, egg parasitism, and a possible substrate-borne alarm signal Revista de Biología Tropical, vol. 64, núm. 3, septiembre, 2016, pp. 1209-1222 Universidad de Costa Rica San Pedro de Montes de Oca, Costa Rica Available in: http://www.redalyc.org/articulo.oa?id=44946472023 How to cite Complete issue Scientific Information System More information about this article Network of Scientific Journals from Latin America, the Caribbean, Spain and Portugal Journal's homepage in redalyc.org Non-profit academic project, developed under the open access initiative Egg-guarding behavior of the treehopper Ennya chrysura (Hemiptera: Membracidae): female aggregations, egg parasitism, and a possible substrate-borne alarm signal Ximena Miranda Escuela de Biología, Universidad de Costa Rica, Ciudad Universitaria Rodrigo Facio, 2060 San José, Costa Rica; [email protected] Received 03-VI-2015. Corrected 09-II-2016. Accepted 09-III-2016. Abstract: Treehoppers are known for their substrate-borne communication and some of them also for their subsocial behavior. Following a more general study of the natural history and substrate-borne signal repertoire of the treehopper Ennya chrysura, the objective of this paper was to explore in greater depth the signals and other behaviors associated specifically to egg-guarding. Theese were studied both in natural and laboratory conditions between July, 2000 and March, 2004. The spatial distribution of egg guarding females was studied in the natural population; recording equipment and playback experiments were used in the laboratory and then analyzed digitally. -
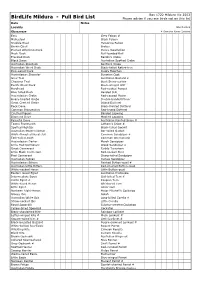
SBOC Full Bird List
Box 1722 Mildura Vic 3502 BirdLife Mildura - Full Bird List Please advise if you see birds not on this list Date Notes Locality March 2012 Observers # Denotes Rarer Species Emu Grey Falcon # Malleefowl Black Falcon Stubble Quail Peregrine Falcon Brown Quail Brolga Plumed Whistling-Duck Purple Swamphen Musk Duck Buff-banded Rail Freckled Duck Baillon's Crake Black Swan Australian Spotted Crake Australian Shelduck Spotless Crake Australian Wood Duck Black-tailed Native-hen Pink-eared Duck Dusky Moorhen Australasian Shoveler Eurasian Coot Grey Teal Australian Bustard # Chestnut Teal Bush Stone-curlew Pacific Black Duck Black-winged Stilt Hardhead Red-necked Avocet Blue-billed Duck Banded Stilt Australasian Grebe Red-capped Plover Hoary-headed Grebe Double-banded Plover Great Crested Grebe Inland Dotterel Rock Dove Black-fronted Dotterel Common Bronzewing Red-kneed Dotterel Crested Pigeon Banded Lapwing Diamond Dove Masked Lapwing Peaceful Dove Australian Painted Snipe # Tawny Frogmouth Latham's Snipe # Spotted Nightjar Black-tailed Godwit Australian Owlet-nightjar Bar-tailed Godwit White-throated Needletail Common Sandpiper # Fork-tailed Swift Common Greenshank Australasian Darter Marsh Sandpiper Little Pied Cormorant Wood Sandpiper # Great Cormorant Ruddy Turnstone Little Black Cormorant Red-necked Stint Pied Cormorant Sharp-tailed Sandpiper Australian Pelican Curlew Sandpiper Australasian Bittern Painted Button-quail # Australian Little Bittern Red-chested Button-quail White-necked Heron Little Button-quail Eastern Great Egret Australian -

Evolution of Bower Complexity and Cerebellum Size in Bowerbirds
Original Paper Brain Behav Evol 2005;66:62–72 Received: September 1, 2004 Returned for revision: September 21, 2004 DOI: 10.1159/000085048 Accepted after revision: January 3, 2005 Published online: April 25, 2005 Evolution of Bower Complexity and Cerebellum Size in Bowerbirds a–c d b Lainy B. Day David A. Westcott Deborah H. Olster a b Departments of Ecology, Evolution, and Marine Biology and Psychology, University of California, c Santa Barbara, Calif. , USA; Department of Zoology and Tropical Ecology, School of Tropical Biology, d James Cook University, Townsville , CSIRO Sustainable Ecosystems and Rainforest CRC, Atherton , Australia Key Words Introduction Birds Bowerbirds Cerebellum Hippocampus Sexual selection Males of the bowerbird family (Ptilonorhynchidae), except three monogamous species, build elaborate dis- play sites (bowers) used to entice females to mate [Mar- Abstract shall, 1954; Kusmierski et al., 1997]. Bower design ap- To entice females to mate, male bowerbirds build elabo- pears to have been sexually selected through female choice rate displays (bowers). Among species, bowers range in as females of several species are known to select mates complexity from simple arenas decorated with leaves to based at least partially on the quality of the bower or the complex twig or grass structures decorated with myriad number of particular items used to decorate the bower colored objects. To investigate the neural underpinnings [Borgia and Mueller, 1992; Madden, 2003a]. Each species of bower building, we examined the contribution of vari- of bowerbird has a particular bower style and preference ation in volume estimates of whole brain (WB), telen- for decorations of certain types or colors [Marshall, 1954; cephalon minus hippocampus (TH), hippocampus (Hp) Madden, 2003b]. -

Function of Note Strings in Japanese Tit Alarm Calls to the Common Cuckoo
Yu et al. Avian Res (2017) 8:22 DOI 10.1186/s40657-017-0080-7 Avian Research RESEARCH Open Access Function of note strings in Japanese Tit alarm calls to the Common Cuckoo: a playback experiment Jiangping Yu1, Weiwei Lv2, Hongwei Xu1, Nehafta Bibi1, Yangyang Yu1, Yunlei Jiang1,3, Wei Liang4 and Haitao Wang1,2* Abstract Background: Birds produce alarm calls to convey information about threats. Some Passerine alarm calls consist of several note strings, but few studies have examined their function. Previous studies have shown that Japanese Tits (Parus minor) can alter the calling rate and number and combination of notes in response to predators. We previously found the combinations of note types in Japanese Tit alarm calls to be signifcantly diferent in response to the Spar- rowhawk (Accipiter nisus) and Common Cuckoo (Cuculus canorus). Methods: Through playback experiments, we tested whether the note strings in Japanese Tit alarm calls to the Com- mon Cuckoo have diferent functions in conveying information. The note strings of selected alarm calls were divided into the categories of C and D, and diferent calls were then constructed separately based on the two note string categories. Original alarm calls (C–D), C calls and D calls were played back to male Japanese Tits during the incubation period. Results: Male Japanese Tits had a signifcantly stronger response to C calls than to C–D calls, and they showed a signifcantly stronger response to both C and C–D calls than to D calls, suggesting that Japanese Tits discriminated between the C and D calls. Conclusions: Our study demonstrated that the C- and D-category note strings of Japanese Tit alarm calls to the Common Cuckoo have diferent functions, which supports the previous fnding that diferent note strings in an alarm call can provide diferent information to receivers.