Sponges from Clipperton Island, East Pacific
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Country of Citizenship Active Exchange Visitors in 2017
Total Number of Active Exchange Visitors by Country of Citizenship in Calendar Year 2017 Active Exchange Visitors Country of Citizenship in 2017 AFGHANISTAN 418 ALBANIA 460 ALGERIA 316 ANDORRA 16 ANGOLA 70 ANTIGUA AND BARBUDA 29 ARGENTINA 8,428 ARMENIA 325 ARUBA 1 ASHMORE AND CARTIER ISLANDS 1 AUSTRALIA 7,133 AUSTRIA 3,278 AZERBAIJAN 434 BAHAMAS, THE 87 BAHRAIN 135 BANGLADESH 514 BARBADOS 58 BASSAS DA INDIA 1 BELARUS 776 BELGIUM 1,938 BELIZE 55 BENIN 61 BERMUDA 14 BHUTAN 63 BOLIVIA 535 BOSNIA AND HERZEGOVINA 728 BOTSWANA 158 BRAZIL 19,231 BRITISH VIRGIN ISLANDS 3 BRUNEI 44 BULGARIA 4,996 BURKINA FASO 79 BURMA 348 BURUNDI 32 CAMBODIA 258 CAMEROON 263 CANADA 9,638 CAPE VERDE 16 CAYMAN ISLANDS 1 CENTRAL AFRICAN REPUBLIC 27 CHAD 32 Total Number of Active Exchange Visitors by Country of Citizenship in Calendar Year 2017 CHILE 3,284 CHINA 70,240 CHRISTMAS ISLAND 2 CLIPPERTON ISLAND 1 COCOS (KEELING) ISLANDS 3 COLOMBIA 9,749 COMOROS 7 CONGO (BRAZZAVILLE) 37 CONGO (KINSHASA) 95 COSTA RICA 1,424 COTE D'IVOIRE 142 CROATIA 1,119 CUBA 140 CYPRUS 175 CZECH REPUBLIC 4,048 DENMARK 3,707 DJIBOUTI 28 DOMINICA 23 DOMINICAN REPUBLIC 4,170 ECUADOR 2,803 EGYPT 2,593 EL SALVADOR 463 EQUATORIAL GUINEA 9 ERITREA 10 ESTONIA 601 ETHIOPIA 395 FIJI 88 FINLAND 1,814 FRANCE 21,242 FRENCH GUIANA 1 FRENCH POLYNESIA 25 GABON 19 GAMBIA, THE 32 GAZA STRIP 104 GEORGIA 555 GERMANY 32,636 GHANA 686 GIBRALTAR 25 GREECE 1,295 GREENLAND 1 GRENADA 60 GUATEMALA 361 GUINEA 40 Total Number of Active Exchange Visitors by Country of Citizenship in Calendar Year 2017 GUINEA‐BISSAU -

New Zealand Oceanographic Institute Memoir 100
ISSN 0083-7903, 100 (Print) ISSN 2538-1016; 100 (Online) , , II COVER PHOTO. Dictyodendrilla cf. cavernosa (Lendenfeld, 1883) (type species of Dictyodendri/la Bergquist, 1980) (see page 24), from NZOI Stn I827, near Rikoriko Cave entrance, Poor Knights Islands Marine Reserve. Photo: Ken Grange, NZOI. This work is licensed under the Creative Commons Attribution-NonCommercial-NoDerivs 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/ NATIONAL INSTITUTE OF WATER AND ATMOSPHERIC RESEARCH The Marine Fauna of New Zealand: Index to the Fauna 2. Porifera by ELLIOT W. DAWSON N .Z. Oceanographic Institute, Wellington New Zealand Oceanographic Institute Memoir 100 1993 • This work is licensed under the Creative Commons Attribution-NonCommercial-NoDerivs 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/ Cataloguing in publication DAWSON, E.W. The marine fauna of New Zealand: Index to the Fauna 2. Porifera / by Elliot W. Dawson - Wellington: New Zealand Oceanographic Institute, 1993. (New Zealand Oceanographic Institute memoir, ISSN 0083-7903, 100) ISBN 0-478-08310-6 I. Title II. Series UDC Series Editor Dennis P. Gordon Typeset by Rose-Marie C. Thompson NIWA Oceanographic (NZOI) National Institute of Water and Atmospheric Research Received for publication: 17 July 1991 © NIWA Copyright 1993 2 This work is licensed under the Creative Commons Attribution-NonCommercial-NoDerivs 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/ CONTENTS Page ABSTRACT 5 INTRODUCTION 5 SCOPE AND ARRANGEMENT 7 SYSTEMATIC LIST 8 Class DEMOSPONGIAE 8 Subclass Homosclcromorpha .............................................................................................. -

Taxonomy and Diversity of the Sponge Fauna from Walters Shoal, a Shallow Seamount in the Western Indian Ocean Region
Taxonomy and diversity of the sponge fauna from Walters Shoal, a shallow seamount in the Western Indian Ocean region By Robyn Pauline Payne A thesis submitted in partial fulfilment of the requirements for the degree of Magister Scientiae in the Department of Biodiversity and Conservation Biology, University of the Western Cape. Supervisors: Dr Toufiek Samaai Prof. Mark J. Gibbons Dr Wayne K. Florence The financial assistance of the National Research Foundation (NRF) towards this research is hereby acknowledged. Opinions expressed and conclusions arrived at, are those of the author and are not necessarily to be attributed to the NRF. December 2015 Taxonomy and diversity of the sponge fauna from Walters Shoal, a shallow seamount in the Western Indian Ocean region Robyn Pauline Payne Keywords Indian Ocean Seamount Walters Shoal Sponges Taxonomy Systematics Diversity Biogeography ii Abstract Taxonomy and diversity of the sponge fauna from Walters Shoal, a shallow seamount in the Western Indian Ocean region R. P. Payne MSc Thesis, Department of Biodiversity and Conservation Biology, University of the Western Cape. Seamounts are poorly understood ubiquitous undersea features, with less than 4% sampled for scientific purposes globally. Consequently, the fauna associated with seamounts in the Indian Ocean remains largely unknown, with less than 300 species recorded. One such feature within this region is Walters Shoal, a shallow seamount located on the South Madagascar Ridge, which is situated approximately 400 nautical miles south of Madagascar and 600 nautical miles east of South Africa. Even though it penetrates the euphotic zone (summit is 15 m below the sea surface) and is protected by the Southern Indian Ocean Deep- Sea Fishers Association, there is a paucity of biodiversity and oceanographic data. -

Appendix: Some Important Early Collections of West Indian Type Specimens, with Historical Notes
Appendix: Some important early collections of West Indian type specimens, with historical notes Duchassaing & Michelotti, 1864 between 1841 and 1864, we gain additional information concerning the sponge memoir, starting with the letter dated 8 May 1855. Jacob Gysbert Samuel van Breda A biography of Placide Duchassaing de Fonbressin was (1788-1867) was professor of botany in Franeker (Hol published by his friend Sagot (1873). Although an aristo land), of botany and zoology in Gent (Belgium), and crat by birth, as we learn from Michelotti's last extant then of zoology and geology in Leyden. Later he went to letter to van Breda, Duchassaing did not add de Fon Haarlem, where he was secretary of the Hollandsche bressin to his name until 1864. Duchassaing was born Maatschappij der Wetenschappen, curator of its cabinet around 1819 on Guadeloupe, in a French-Creole family of natural history, and director of Teyler's Museum of of planters. He was sent to school in Paris, first to the minerals, fossils and physical instruments. Van Breda Lycee Louis-le-Grand, then to University. He finished traveled extensively in Europe collecting fossils, especial his studies in 1844 with a doctorate in medicine and two ly in Italy. Michelotti exchanged collections of fossils additional theses in geology and zoology. He then settled with him over a long period of time, and was received as on Guadeloupe as physician. Because of social unrest foreign member of the Hollandsche Maatschappij der after the freeing of native labor, he left Guadeloupe W etenschappen in 1842. The two chief papers of Miche around 1848, and visited several islands of the Antilles lotti on fossils were published by the Hollandsche Maat (notably Nevis, Sint Eustatius, St. -
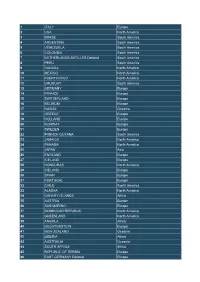
1 ITALY Europe 2 USA North America 3 BRASIL South America 4
1 ITALY Europe 2 USA North America 3 BRASIL South America 4 ARGENTINA South America 5 VENEZUELA South America 6 COLOMBIA South America 7 NETHERLANDS ANTILLES Deleted South America 8 PERU South America 9 CANADA North America 10 MEXICO North America 11 PUERTO RICO North America 12 URUGUAY South America 13 GERMANY Europe 14 FRANCE Europe 15 SWITZERLAND Europe 16 BELGIUM Europe 17 HAWAII Oceania 18 GREECE Europe 19 HOLLAND Europe 20 NORWAY Europe 21 SWEDEN Europe 22 FRENCH GUYANA South America 23 JAMAICA North America 24 PANAMA North America 25 JAPAN Asia 26 ENGLAND Europe 27 ICELAND Europe 28 HONDURAS North America 29 IRELAND Europe 30 SPAIN Europe 31 PORTUGAL Europe 32 CHILE South America 33 ALASKA North America 34 CANARY ISLANDS Africa 35 AUSTRIA Europe 36 SAN MARINO Europe 37 DOMINICAN REPUBLIC North America 38 GREENLAND North America 39 ANGOLA Africa 40 LIECHTENSTEIN Europe 41 NEW ZEALAND Oceania 42 LIBERIA Africa 43 AUSTRALIA Oceania 44 SOUTH AFRICA Africa 45 REPUBLIC OF SERBIA Europe 46 EAST GERMANY Deleted Europe 47 DENMARK Europe 48 SAUDI ARABIA Asia 49 BALEARIC ISLANDS Europe 50 RUSSIA Europe 51 ANDORA Europe 52 FAROER ISLANDS Europe 53 EL SALVADOR North America 54 LUXEMBOURG Europe 55 GIBRALTAR Europe 56 FINLAND Europe 57 INDIA Asia 58 EAST MALAYSIA Oceania 59 DODECANESE ISLANDS Europe 60 HONG KONG Asia 61 ECUADOR South America 62 GUAM ISLAND Oceania 63 ST HELENA ISLAND Africa 64 SENEGAL Africa 65 SIERRA LEONE Africa 66 MAURITANIA Africa 67 PARAGUAY South America 68 NORTHERN IRELAND Europe 69 COSTA RICA North America 70 AMERICAN -

California Marine Life Protection Act (MLPA) Initiative
California Marine Life Protection Act (MLPA) Initiative Evaluation of Existing Central Coast Marine Protected Areas DRAFT, v. 2.0 November 4, 2005 CONTENTS EXECUTIVE SUMMARY ..........................................................................................................................4 1.0 INTRODUCTION ................................................................................................................................8 2.0 EVALUATION OF EXISTING MPAs .................................................................................................11 2.1 Año Nuevo Special Closure...........................................................................................................11 2.2 Elkhorn Slough State Marine Reserve ...........................................................................................12 2.3 Hopkins State Marine Reserve......................................................................................................15 2.4 Pacific Grove State Marine Conservation Area.............................................................................16 2.5: Carmel Bay State Marine Conservation Area ..............................................................................17 2.6 Point Lobos State Marine Reserve................................................................................................19 2.7 Julia Pfeiffer Burns State Marine Conservation Area.....................................................................20 2.8 Big Creek State Marine Reserve....................................................................................................22 -

The Political, Security, and Climate Landscape in Oceania
The Political, Security, and Climate Landscape in Oceania Prepared for the US Department of Defense’s Center for Excellence in Disaster Management and Humanitarian Assistance May 2020 Written by: Jonah Bhide Grace Frazor Charlotte Gorman Claire Huitt Christopher Zimmer Under the supervision of Dr. Joshua Busby 2 Table of Contents Executive Summary 3 United States 8 Oceania 22 China 30 Australia 41 New Zealand 48 France 53 Japan 61 Policy Recommendations for US Government 66 3 Executive Summary Research Question The current strategic landscape in Oceania comprises a variety of complex and cross-cutting themes. The most salient of which is climate change and its impact on multilateral political networks, the security and resilience of governments, sustainable development, and geopolitical competition. These challenges pose both opportunities and threats to each regionally-invested government, including the United States — a power present in the region since the Second World War. This report sets out to answer the following questions: what are the current state of international affairs, complexities, risks, and potential opportunities regarding climate security issues and geostrategic competition in Oceania? And, what policy recommendations and approaches should the US government explore to improve its regional standing and secure its national interests? The report serves as a primer to explain and analyze the region’s state of affairs, and to discuss possible ways forward for the US government. Given that we conducted research from August 2019 through May 2020, the global health crisis caused by the novel coronavirus added additional challenges like cancelling fieldwork travel. However, the pandemic has factored into some of the analysis in this report to offer a first look at what new opportunities and perils the United States will face in this space. -

Proposal for a Revised Classification of the Demospongiae (Porifera) Christine Morrow1 and Paco Cárdenas2,3*
Morrow and Cárdenas Frontiers in Zoology (2015) 12:7 DOI 10.1186/s12983-015-0099-8 DEBATE Open Access Proposal for a revised classification of the Demospongiae (Porifera) Christine Morrow1 and Paco Cárdenas2,3* Abstract Background: Demospongiae is the largest sponge class including 81% of all living sponges with nearly 7,000 species worldwide. Systema Porifera (2002) was the result of a large international collaboration to update the Demospongiae higher taxa classification, essentially based on morphological data. Since then, an increasing number of molecular phylogenetic studies have considerably shaken this taxonomic framework, with numerous polyphyletic groups revealed or confirmed and new clades discovered. And yet, despite a few taxonomical changes, the overall framework of the Systema Porifera classification still stands and is used as it is by the scientific community. This has led to a widening phylogeny/classification gap which creates biases and inconsistencies for the many end-users of this classification and ultimately impedes our understanding of today’s marine ecosystems and evolutionary processes. In an attempt to bridge this phylogeny/classification gap, we propose to officially revise the higher taxa Demospongiae classification. Discussion: We propose a revision of the Demospongiae higher taxa classification, essentially based on molecular data of the last ten years. We recommend the use of three subclasses: Verongimorpha, Keratosa and Heteroscleromorpha. We retain seven (Agelasida, Chondrosiida, Dendroceratida, Dictyoceratida, Haplosclerida, Poecilosclerida, Verongiida) of the 13 orders from Systema Porifera. We recommend the abandonment of five order names (Hadromerida, Halichondrida, Halisarcida, lithistids, Verticillitida) and resurrect or upgrade six order names (Axinellida, Merliida, Spongillida, Sphaerocladina, Suberitida, Tetractinellida). Finally, we create seven new orders (Bubarida, Desmacellida, Polymastiida, Scopalinida, Clionaida, Tethyida, Trachycladida). -

Carnivorous Sponges of the Atlantic and Arctic Oceans
&DUQLYRURXVVSRQJHVRIWKH$WODQWLFDQG $UFWLF2FHDQV 3K\ORJHQ\WD[RQRP\GLVWULEXWLRQDQGPLFURELDODVVRFLDWLRQVRIWKH &ODGRUKL]LGDH 'HPRVSRQJLDH3RHFLORVFOHULGD -RQ7KRPDVVHQ+HVWHWXQ Dissertation for the degree of philosophiae doctor (PhD) at the University of Bergen 'LVVHUWDWLRQGDWH1RYHPEHUWK © Copyright Jon Thomassen Hestetun The material in this publication is protected by copyright law. Year: 2016 Title: Carnivorous sponges of the Atlantic and Arctic Oceans Phylogeny, taxonomy, distribution and microbial associations of the Cladorhizidae (Demospongiae, Poecilosclerida) Author: Jon Thomassen Hestetun Print: AiT Bjerch AS / University of Bergen 3 Scientific environment This PhD project was financed through a four-year PhD position at the University of Bergen, and the study was conducted at the Department of Biology, Marine biodiversity research group, and the Centre of Excellence (SFF) Centre for Geobiology at the University of Bergen. The work was additionally funded by grants from the Norwegian Biodiversity Centre (grant to H.T. Rapp, project number 70184219), the Norwegian Academy of Science and Letters (grant to H.T. Rapp), the Research Council of Norway (through contract number 179560), the SponGES project through Horizon 2020, the European Union Framework Programme for Research and Innovation (grant agreement No 679849), the Meltzer Fund, and the Joint Fund for the Advancement of Biological Research at the University of Bergen. 4 5 Acknowledgements I have, initially through my master’s thesis and now during these four years of my PhD, in all been involved with carnivorous sponges for some six years. Trying to look back and somehow summarizing my experience with this work a certain realization springs to mind: It took some time before I understood my luck. My first in-depth exposure to sponges was in undergraduate zoology, and I especially remember watching “The Shape of Life”, an American PBS-produced documentary series focusing on the different animal phyla, with an enthusiastic Dr. -

Revision of the Genus Trachytedania (Porifera: Poecilosclerida) with A
J. Mar. Biol. Ass. U.K. 2001), 81,569^579 Printed in the United Kingdom Revision of the genus Trachytedania Porifera: Poecilosclerida) with a description of Trachytedania ferrolensis sp. nov. from the north-east Atlantic F.J. Cristobo*and V. Urgorri O *Departamento de Biodiversidade e Recursos Marin¬ os, Instituto de Acuicultura, Universidade de Santiago de Compostela, 15706 Santiago de Compostela, Spain. E-mail: [email protected] OLaboratorio de Zoolox|¨a Marin¬ a, Departamento de Biolox|¨a Animal, Universidade de Santiago de Compostela, 15706 Santiago de Compostela, Spain. E-mail: [email protected] The genus Trachytedania Ridley, 1881 Porifera: Poecilosclerida), includes those tedaniidae sponges characterized by the presence of acanthostyles. To date, only ¢ve species of the genus Trachytedania have been described of which only three actually belong to this genus. Other species described as Tedania could be considered as Trachytedania because of the presence of acanthostyles in their skeletons. Some authors propose retaining Trachytedania as a subgenus of Tedania because of its skeletal structure. In this work, a taxonomic revision and consideration of the validity of the genus is provided. The morphology and anatomy of Trachytedania ferrolensis sp. nov. from the R|¨a de Ferrol Spain, north-east Atlantic) are described in detail, and the types of the three valid species described as Trachytedania to date: Trachytedania spinata, Trachytedania patagonica and Trachytedania biraphidora are studied. A key to this genus is provided. INTRODUCTION MATERIALS AND METHODS The position of the Family Tedaniidae within the The specimens were collected by SCUBA divers and by Poecilosclerida is still disputed. Species of the genus dredging using a naturalist benthic dredge Holme & Tedania Gray, 1867, have always been di¤cult to di¡er- McIntyre, 1984) in the sublittoral zones of R|¨a de Ferrol entiate clearly Bergquist & Fromont, 1988) and the value Spain). -

Porifera) in Singapore and Description of a New Species of Forcepia (Poecilosclerida: Coelosphaeridae)
Contributions to Zoology, 81 (1) 55-71 (2012) Biodiversity of shallow-water sponges (Porifera) in Singapore and description of a new species of Forcepia (Poecilosclerida: Coelosphaeridae) Swee-Cheng Lim1, 3, Nicole J. de Voogd2, Koh-Siang Tan1 1 Tropical Marine Science Institute, National University of Singapore, 18 Kent Ridge Road, Singapore 119227, Singapore 2 Netherlands Centre for Biodiversity, Naturalis, PO Box 9517, 2300 RA Leiden, The Netherlands 3 E-mail: [email protected] Key words: intertidal, Southeast Asia, sponge assemblage, subtidal, tropical Abstract gia) patera (Hardwicke, 1822) was the first sponge de- scribed from Singapore in the 19th century. This was A surprisingly high number of shallow water sponge species followed by Leucosolenia flexilis (Haeckel, 1872), (197) were recorded from extensive sampling of natural inter- Coelocarteria singaporensis (Carter, 1883) (as Phloeo tidal and subtidal habitats in Singapore (Southeast Asia) from May 2003 to June 2010. This is in spite of a highly modified dictyon), and Callyspongia (Cladochalina) diffusa coastline that encompasses one of the world’s largest container Ridley (1884). Subsequently, Dragnewitsch (1906) re- ports as well as extensive oil refining and bunkering industries. corded 24 sponge species from Tanjong Pagar and Pu- A total of 99 intertidal species was recorded in this study. Of lau Brani in the Singapore Strait. A further six species these, 53 species were recorded exclusively from the intertidal of sponge were reported from Singapore in the 1900s, zone and only 45 species were found on both intertidal and subtidal habitats, suggesting that tropical intertidal and subtidal although two species, namely Cinachyrella globulosa sponge assemblages are different and distinct. -

From Northern Bass Strait, Southern Australia
31 August 1989 Memoirs of the Museum of Victoria 50(1): 1-242 (1989) ISSN 0814-1827 https://doi.org/10.24199/j.mmv.1989.50.01 DEMOSPONGIAE (PORIFERA) FROM NORTHERN BASS STRAIT, SOUTHERN AUSTRALIA By Felix Wiedenmayer Department of Invertebrate Zoology, Museum of Victoria, Swanston Street, Melbourne, Victoria 3000, Australia Present address: Naturhistorisches Museum Basel, Agustinergasse 2, 4001 Basel, Switzerland Abstract Wiedenmayer, F., 1989. Demospongiae from northern Bass Strait, southern Australia. Memoirs of the Museum of Victoria 50(1): 1-242. Eighty-four species (in 47 genera) in the Museum of Victoria, Melbourne, are described and illustrated. Of these, 21 species are described as new: Ancorina repens, A. suina, Stelletta arenitecta, Rhabdastrella cordata, R. intermedia, Tetilla praecipua, Latrunculia hallmanni, Pseudaxinella decipiens, Reniochalina sectilis, Rhaphoxya felina, Clathria wilsoni, Echinoclathria egena, Psammoclema bitextum, P. fissuratum, P. goniodes, P. radiatum, P. stipitatum, P. van- soesti, Callyspongia persculpta, C. toxifera, and Thorecta glomerosus. Eighteen records are new for the Maugean province, and three (Phorbas tenacior, Darwinella gardineri, and Gel- liodes incrustans) are new for the Australian fauna. The following revisions depart from those adopted in Wiedenmayer et al. (in press). The family Desmacididae is divided into Desmacidi- nae and Stylotellinae, and the genera Stylotella ( = Batzella), Phoriospongia ( = Chondropsis), and Psammoclema ( = Psammopemma, Sarcocornea) are assigned to the latter. Dactylia, Chalinopsilla and Arenosclera are synonymised with Callyspongia. Thorectandra is synonymised with Thorecta. Dendrilla cactos (Selenka) is a senior synonym of D. rosea Lendenfeld. The composition of this collection is even, with respect to the known demosponge fauna of Victoria and Tasmania. Its zoogeographic affinity is essentially Indo-West Pacific and relictic Tethyan, its provincial endemism high, and its overlap with the Antarctic/Subantarctic fauna almost nil.