Name Genbank
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Anti-Inflammatory Effects of V-3 Polyunsaturated Fatty Acids And
ORIGINAL ARTICLE Anti-inflammatory Effects of v-3 Polyunsaturated Fatty Acids and Soluble Epoxide Hydrolase Inhibitors in Angiotensin-II–Dependent Hypertension Arzu Ulu, MSc,* Todd R. Harris, PhD,* Christophe Morisseau, PhD,* Christina Miyabe, BS,* Hiromi Inoue, DVM,† Gertrud Schuster, PhD,‡ Hua Dong, PhD,* Ana-Maria Iosif, PhD,§ Jun-Yan Liu, PhD,* Robert H. Weiss, MD,†¶ Nipavan Chiamvimonvat, MD,k¶ John D. Imig, PhD,** and Bruce D. Hammock, PhD* renal markers of inflammation (ie, prostaglandins and MCP-1), Abstract: The mechanisms underlying the anti-inflammatory and downregulated an epithelial sodium channel, and upregulated angioten- antihypertensive effects of long-chain v-3 polyunsaturated fatty acids sin-converting enzyme-2 message and significantly modulated cyclo- (v-3 PUFAs) are still unclear. The epoxides of an v-6 fatty acid, oxygenase and lipoxygenase metabolic pathways. Overall, our findings arachidonic acid epoxyeicosatrienoic acids also exhibit antihyperten- suggest that epoxides of the v-3 PUFAs contribute to lowering systolic sive and anti-inflammatory effects. Thus, we hypothesized that the blood pressure and attenuating inflammation in part by reduced pros- major v-3 PUFAs, including eicosapentaenoic acid (EPA) and doco- taglandinsandMCP-1andbyupregulation of angiotensin-converting sahexaenoic acid (DHA), may lower the blood pressure and attenuate enzyme-2 in angiotensin-II–dependent hypertension. renal markers of inflammation through their epoxide metabolites. v Here, we supplemented mice with an v-3 rich diet for 3 weeks in Key Words: -3 polyunsaturated fatty acids, EPA, DHA, soluble – a murine model of angiotensin-II–dependent hypertension. Also, epoxide hydrolase inhibitors, angiotensin-II dependent hypertension because EPA and DHA epoxides are metabolized by soluble epoxide (J Cardiovasc PharmacolÔ 2013;62:285–297) hydrolase (sEH), we tested the combination of an sEH inhibitor and the v-3 rich diet. -

Gene Symbol Gene Description ACVR1B Activin a Receptor, Type IB
Table S1. Kinase clones included in human kinase cDNA library for yeast two-hybrid screening Gene Symbol Gene Description ACVR1B activin A receptor, type IB ADCK2 aarF domain containing kinase 2 ADCK4 aarF domain containing kinase 4 AGK multiple substrate lipid kinase;MULK AK1 adenylate kinase 1 AK3 adenylate kinase 3 like 1 AK3L1 adenylate kinase 3 ALDH18A1 aldehyde dehydrogenase 18 family, member A1;ALDH18A1 ALK anaplastic lymphoma kinase (Ki-1) ALPK1 alpha-kinase 1 ALPK2 alpha-kinase 2 AMHR2 anti-Mullerian hormone receptor, type II ARAF v-raf murine sarcoma 3611 viral oncogene homolog 1 ARSG arylsulfatase G;ARSG AURKB aurora kinase B AURKC aurora kinase C BCKDK branched chain alpha-ketoacid dehydrogenase kinase BMPR1A bone morphogenetic protein receptor, type IA BMPR2 bone morphogenetic protein receptor, type II (serine/threonine kinase) BRAF v-raf murine sarcoma viral oncogene homolog B1 BRD3 bromodomain containing 3 BRD4 bromodomain containing 4 BTK Bruton agammaglobulinemia tyrosine kinase BUB1 BUB1 budding uninhibited by benzimidazoles 1 homolog (yeast) BUB1B BUB1 budding uninhibited by benzimidazoles 1 homolog beta (yeast) C9orf98 chromosome 9 open reading frame 98;C9orf98 CABC1 chaperone, ABC1 activity of bc1 complex like (S. pombe) CALM1 calmodulin 1 (phosphorylase kinase, delta) CALM2 calmodulin 2 (phosphorylase kinase, delta) CALM3 calmodulin 3 (phosphorylase kinase, delta) CAMK1 calcium/calmodulin-dependent protein kinase I CAMK2A calcium/calmodulin-dependent protein kinase (CaM kinase) II alpha CAMK2B calcium/calmodulin-dependent -

Uvic Thesis Template
Characterization of A-Type Ephrin Signaling by Jessa Bazowski B.Sc, University of Victoria, 2004 A Thesis Submitted in Partial Fulfillment of the Requirements for the Degree of MASTER OF SCIENCE in the Department of Biology Jessa Bazowski, 2007 University of Victoria All rights reserved. This thesis may not be reproduced in whole or in part, by photocopy or other means, without the permission of the author. ii Characterization of A-Type Ephrin Signaling by Jessa Bazowski B.Sc, University of Victoria, 2004 Supervisory Committee Dr. Perry L. Howard (Department of Biology) Supervisor Dr. Robert Ingham (Department of Biology) Co-Supervisor or Departmental Member Dr. Robert Burke (Department of Biology) Departmental Member Dr. Caroline Cameron (Department of Biochemistry and Microbiology) Outside Member iii Abstract Supervisory Committee Dr. Perry L. Howard (Department of Biology) Supervisor Dr. Robert Ingham (Department of Biology) Co-Supervisor or Departmental Member Dr. Robert Burke (Department of Biology) Departmental Member Dr. Caroline Cameron (Department of Biochemistry and Microbiology) Outside Member Membrane attachment of ephrin ligands plays an important role in Eph receptor activation. Membrane anchorage is thought to provide a clustering effect to ephrins that is necessary for stimulation of Eph receptor kinase activity. The presence of soluble A-type ephrin in conditioned media of numerous cultured cancer cell lines and normal endothelial cells prompted me to question the purpose of ephrin release. In this thesis I show that ephrin A1, a potent angiogenic factor, is released from several cancer cell lines and is a substrate for tissue transglutaminase, a multifunctional enzyme with the ability to form covalent crosslinks between substrate proteins. -

Airway Smooth Muscle Cells Are Insensitive to the Anti-Proliferative
Immunobiology 224 (2019) 490–496 Contents lists available at ScienceDirect Immunobiology journal homepage: www.elsevier.com/locate/imbio Airway smooth muscle cells are insensitive to the anti-proliferative effects of corticosteroids: The novel role of insulin growth factor binding Protein-1 in T asthma ⁎ Hong Buia, Yassine Amranib, Brian Deeneyc, Reynold A. Panettieric, Omar Tlibad, a Penn Health System, Microbiology laboratories, University of Pennsylvania, Philadelphia, PA, USA b Institute for Lung Health, Department of Infection, Inflammation and Immunity, University of Leicester, United Kingdom c Rutgers Institute of Translational Medicine and Sciences, Rutgers School of Medicine, New Brunswick, NJ, USA d Department of Biomedical Sciences, College of Veterinary Medicine, Long Island University, Brookville, NY, USA ARTICLE INFO ABSTRACT Keywords: Airway remodeling in asthma manifests, in part, as enhanced airway smooth muscle (ASM) mass, due to myocyte Asthma proliferation. While the anti-proliferative effects of glucocorticoid (GC) were investigated in normal ASM cells Glucocorticoid insensitivity (NASMC), little is known about such effects in ASM cells derived from asthma subjects (AASMC). We posit that Airway smooth muscle GC differentially modulates mitogen-induced proliferation of AASMC and NASMC. Cells were cultured, starved, Proliferation then treated with Epidermal growth factor (EGF) (10 ng/ml) and Platelet-derived growth factor (PDGF) (10 ng/ Growth ml) for 24 h and/or fluticasone propionate (FP) (100 nM) added 2 h before. Cell counts and flow cytometry Airway remodeling analyses showed that FP failed to decrease the cell number of and DNA synthesis in AASMC irrespective of mitogens used. We also examine the ability of Insulin Growth Factor Binding Protein-1 (IGFBP-1), a steroid- inducible gene that deters cell growth in other cell types, to inhibit proliferation of AASMC where FP failed. -

Epigenetic Loss of the RNA Decapping Enzyme NUDT16 Mediates C-MYC Activation in T-Cell Acute Lymphoblastic Leukemia
OPEN Leukemia (2017) 31, 1622–1657 www.nature.com/leu LETTERS TO THE EDITOR Epigenetic loss of the RNA decapping enzyme NUDT16 mediates C-MYC activation in T-cell acute lymphoblastic leukemia Leukemia (2017) 31, 1622–1625; doi:10.1038/leu.2017.99 Having found the aforementioned NUDT16 CpG island methy- lation profiles, we studied in greater detail their association with the possible transcriptional inactivation of the NUDT16 gene at the RNA and protein levels in leukemia cell lines. We first It is possible that the occurrence of intrinsic defects in RNA performed bisulfite genomic sequencing of mutiple clones in the – processing pathways, such as RNA decapping,1 3 contribute to the T-cell Acute Lymphoblastic Leukemia (T-ALL) cell lines CCRF-CEM, distorted RNA landscapes of cancer cells. After transcription by Jurkat, MOLT-4 and MOLT-16 using primers that encompassed the RNA polymerase II, RNA molecules are equipped with a 5´-end transcription start site-associated CpG island and confirmed the N7-methyl guanosine (m7G)-cap. This m7G-cap is essential for hypermethylated status of the 5′-end region of NUDT16 in translation, stabilizing the RNA molecule and protecting it from comparison to normal T lymphocytes (Figure 1b), validating the – exonucleolytic breakdown.1 3 For RNA decay to occur the DNA methylation patterns obtained by the microarray approach m7G-cap first needs to be removed. This process is known as (Supplementary Figure S2). In contrast, normal T lymphocytes, the – decapping.1 3 The decapping mRNA 2 (DCP2) enzyme,4 also T-ALL cell lines KOPN-8, REH and RS4;11 and the leukemia cell known as the nucleoside diphosphate-linked moiety X motif lines HL-60 and K562 derived from myeloid lineage were all found 20 (NUDT20), was originally thought to be the only mammalian to be unmethylated (Figure 1b; Supplementary Figure S2). -

CDH12 Cadherin 12, Type 2 N-Cadherin 2 RPL5 Ribosomal
5 6 6 5 . 4 2 1 1 1 2 4 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 2 2 A A A A A A A A A A A A A A A A A A A A C C C C C C C C C C C C C C C C C C C C R R R R R R R R R R R R R R R R R R R R B , B B B B B B B B B B B B B B B B B B B , 9 , , , , 4 , , 3 0 , , , , , , , , 6 2 , , 5 , 0 8 6 4 , 7 5 7 0 2 8 9 1 3 3 3 1 1 7 5 0 4 1 4 0 7 1 0 2 0 6 7 8 0 2 5 7 8 0 3 8 5 4 9 0 1 0 8 8 3 5 6 7 4 7 9 5 2 1 1 8 2 2 1 7 9 6 2 1 7 1 1 0 4 5 3 5 8 9 1 0 0 4 2 5 0 8 1 4 1 6 9 0 0 6 3 6 9 1 0 9 0 3 8 1 3 5 6 3 6 0 4 2 6 1 0 1 2 1 9 9 7 9 5 7 1 5 8 9 8 8 2 1 9 9 1 1 1 9 6 9 8 9 7 8 4 5 8 8 6 4 8 1 1 2 8 6 2 7 9 8 3 5 4 3 2 1 7 9 5 3 1 3 2 1 2 9 5 1 1 1 1 1 1 5 9 5 3 2 6 3 4 1 3 1 1 4 1 4 1 7 1 3 4 3 2 7 6 4 2 7 2 1 2 1 5 1 6 3 5 6 1 3 6 4 7 1 6 5 1 1 4 1 6 1 7 6 4 7 e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e e l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m -

IGFBP5) Reverses Cisplatin-Resistance in Esophageal Carcinoma
cells Article Expression of Insulin-Like Growth Factor Binding Protein-5 (IGFBP5) Reverses Cisplatin-Resistance in Esophageal Carcinoma Dessy Chan 1,†, Yuanyuan Zhou 1,†, Chung Hin Chui 1, Kim Hung Lam 1, Simon Law 2, Albert Sun-chi Chan 3, Xingshu Li 3,*, Alfred King-yin Lam 4,* and Johnny Cheuk On Tang 1,* 1 State Key Laboratory of Chemical Biology and Drug Discovery, Lo Ka Chung Centre for Natural Anti-cancer Drug Development, Department of Applied Biology and Chemical Technology, The Hong Kong Polytechnic University, Hong Kong, China; [email protected] (D.C.); [email protected] (Y.Z.); [email protected] (C.H.C.), [email protected] (K.H.L.) 2 Department of Surgery, Li Ka Shing Faculty of Medicine, The University of Hong Kong, Hong Kong, China; [email protected] 3 School of Pharmaceutical Sciences, Sun Yat-sen University, Guangzhou 510006, China; [email protected] 4 Griffith Medical School, Griffith University, Gold Coast, QLD 4222, Australia * Correspondence: [email protected] (X.L.); A.Lam@griffith.edu.au (A.K.L.); [email protected] (J.C.O.T.); Tel.: +852-3400-8727 (J.C.O.T.) † These authors contributed equally to this work. Received: 3 September 2018; Accepted: 16 September 2018; Published: 20 September 2018 Abstract: Cisplatin (CDDP) is one of the front-line chemotherapeutic drugs used in the treatment of esophageal squamous cell carcinoma (ESCC). Occurrence of resistance to CDDP has become one of the main challenges in cancer therapy. In this study, the gene expression profile of CDDP-resistant ESCC cells was investigated and molecular approaches were explored in an attempt to reverse the CDDP resistance. -

Contents Supplemental Table 1
Supplementary material Open Heart SUPPLEMENTAL MATERIAL TO “EXPLORATION OF PATHOPHYSIOLOGICAL PATHWAYS FOR INCIDENT ATRIAL FIBRILLATION – THE MALMÖ PREVENTIVE PROJECT” John Molvin, Amra Jujic, Olle Melander, Manan Pareek, Lennart Råstam, Ulf Lindblad, Bledar Daka, Margrét Leósdóttir, Peter M. Nilsson, Michael H. Olsen & Martin Magnusson Contents Supplemental table 1. Unadjusted Cox regression analyses examining all 92 proteins relation to incident atrial fibrillation ................................................................................... 2-3 List of abbreviations……………………………………………………………………………………………………………4 Molvin J, et al. Open Heart 2020; 7:e001190. doi: 10.1136/openhrt-2019-001190 Supplementary material Open Heart Supplemental table 1. Unadjusted Cox regression analyses examining all 92 proteins relation to incident atrial fibrillation Protein Hazard ratio (95% confidence interval) p-value PON3 0.80 (0.72-0.89) 7.3x10-5 IGFBP2 4.47 (1.42-14-1) 0.011 PAI 1.44 (0.65-3.18) 0.371 CTSD 2.45 (1.13-5.30) 0.023 FABP4 1.27 (1.13-1.44) 8.6x10-5 CD163 5.25 (1.14-24.1) 0.033 GAL4 1.30 (1.15-1.47) 3.5x10-5 LDL-receptor 0.81 (0.39-1.69) 0.582 IL1RT2 0.75 (0.24-2.34) 0.614 t-PA 2.75 (1.21-6.27) 0.016 SELE 0.99 (0.51-1.90) 0.969 CTSZ 2.97 (1.00-8.78) 0.050 GDF15 1.41 (1.25-1.59) 9.7x10-9 CSTB 3.75 (1.58-8.92) 0.003 MPO 4.48 (1.73-11.7) 0.002 PCSK9 1.18 (0.73-1.93) 0.501 IGFBP1 2.48 (1.42-4.35) 0.001 RARRES2 64.3 (1.87-2220.8) 0.021 ITGB2 1.01 (0.31-3.29) 0.990 CCL15 3.58 (0.96-13.3) 0.057 SCGB3A2 0.97 (0.71-1.32) 0.839 CHI3L1 1.26 (1.12-1.43) -

Inhibiting the Soluble Epoxide Hydrolase Increases the Epfas and ERK1/2 Expression in the Hippocampus of Licl-Pilocarpine Post-Status Epilepticus Rat Model
Inhibiting the soluble epoxide hydrolase increases the EpFAs and ERK1/2 expression in the hippocampus of LiCl-pilocarpine post-status epilepticus rat model Weifeng Peng Zhongshan Hospital Fudan University https://orcid.org/0000-0002-6225-1409 Yijun Shen Zhongshan Hospital Fudan University Qiang Wang Zhongshan Hospital Fudan University Jing Ding ( [email protected] ) Zhongshan Hospital Fudan University Xin Wang ( [email protected] ) Zhongshan Hospital Fudan University Research Article Keywords: epilepsy, soluble epoxide hydrolase, epoxygenated fatty acids (EpFAs), extracellular signal- activated protein kinase 1/2 (ERK1/2) Posted Date: April 15th, 2021 DOI: https://doi.org/10.21203/rs.3.rs-385335/v1 License: This work is licensed under a Creative Commons Attribution 4.0 International License. Read Full License Page 1/20 Abstract Purpose This study aimed to investigate the enzyme activity of soluble epoxide hydrolase (sEH) and quantify metabolic substrates i.e. epoxygenated fatty acids (EpFAs) and products of sEH in the hippocampus after administering TPPU [1-triuoromethoxyphenyl-3-(1-propionylpiperidin-4-yl)urea], the inhibitor of sEH, and further explored whether the extracellular signal-activated protein kinase 1/2 (ERK1/2) was involved in the anti-seizure effect of TPPU in the lithium chloride (LiCl)-pilocarpine induced post-status epilepticus (SE) rat model. Methods The rats were intraperitoneally (I.P.) injected with LiCl and pilocarpine to induce SE and then spontaneous recurrent seizures (SRS) were observed. Rats were randomly assigned into SRS + 0.1 TPPU group (intragastrically administering 0.1 mg/kg/d TPPU), SRS + PEG 400 group (administering the vehicle instead), and Control group. -

Regulation of the Bovine Oviductal Fluid Proteome
REPRODUCTIONRESEARCH Regulation of the bovine oviductal fluid proteome Julie Lamy1, Valérie Labas1,2, Grégoire Harichaux1,2, Guillaume Tsikis1, Pascal Mermillod1 and Marie Saint-Dizier1,3 1Physiologie de la Reproduction et des Comportements (PRC), INRA, CNRS, IFCE, Université de Tours, Nouzilly, France, 2INRA, Plateforme d’Analyse Intégrative des Biomolécules (PAIB), Nouzilly, France and 3Université François Rabelais de Tours, UFR Sciences et Techniques, Tours, France Correspondence should be addressed to M Saint-Dizier; Email: [email protected] Abstract Our objective was to investigate the regulation of the proteome in the bovine oviductal fluid according to the stage of the oestrous cycle, to the side relative to ovulation and to local concentrations of steroid hormones. Luminal fluid samples from both oviducts were collected at four stages of the oestrous cycle: pre-ovulatory (Pre-ov), post-ovulatory (Post-ov), and mid- and late luteal phases from adult cyclic cows (18–25 cows/stage). The proteomes were assessed by nanoLC–MS/MS and quantified by label-free method. Totally, 482 proteins were identified including a limited number of proteins specific to one stage or one side. Proportions of differentially abundant proteins fluctuated from 10 to 24% between sides at one stage and from 4 to 20% among stages in a given side of ovulation. In oviductal fluids ipsilateral to ovulation, Annexin A1 was the most abundant protein at Pre-ov compared with Post-ov while numerous heat shock proteins were more abundant at Post-ov compared with Pre-ov. Among differentially abundant proteins, seven tended to be correlated with intra-oviductal concentrations of progesterone. -
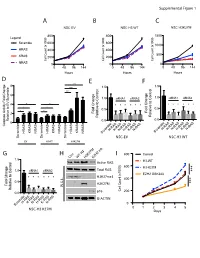
C a B D G E F
Supplemental Figure 1 A B C NSC EV NSC H3 WT NSC H3K27M 800 800 1500 ) ) Legend ) Scramble 600 600 x1000 x1000 x1000 ( ( ( 1000 t t t n n HRAS n 400 400 u u u o o o C C C 500 l l KRAS l 200 200 el el el C C C NRAS 0 0 0 0 48 96 144 0 48 96 144 0 48 96 144 Hours Hours Hours D *** E F 8 1.5 1.5 e *** l l *** ro ro t 6 t 1.0 siRNA1 siRNA2 1.0 siRNA1 siRNA2 Con Con o o t 4 * * t * * * * * * e * * * * * * e v i * * v i t 0.5 * * t 0.5 Fold Change 2 Fold Change Rela Rela Relative to EV Scrambl 0.0 0.0 0 Caspase Activity Fold Change H-RASK-RASN-RASH-RASK-RASN-RAS H-RASK-RASN-RASH-RASK-RASN-RAS KRAS KRAS KRAS HRAS NRAS HRAS NRAS HRAS NRAS Scramble Scramble Scramble Scramble Scramble NSC-EV NSC-H3 WT EV H3WT H3K27M G H I 800 Control Con WT H3 H3K27MEZH2 inb 1.5 l H3-WT Active RAS ro ) t 600 H3-K27M 1.0 siRNA1 siRNA2 Total RAS *** Con x1000 ( o EZH2 GSK343 t t * * * * * * H3K27me3 n e 400 u v i o t 0.5 *** H3K27M C Fold Change l W.C.L. el Rela p16 C 200 0.0 B-ACTIN H-RASK-RASN-RASH-RASK-RASN-RAS 0 Scramble NSC-H3 K27M 0 1 2 3 4 5 Days E C A 100 Cell Viability 100 25 50 75 Cell Viability 25 50 75 0 0 Fold Change MOCK MOCK Relative to Control Scramble * 0.0 0.2 0.4 0.6 0.8 1.0 1.2 MYC Scramble MYC MYC * PDGFRA Scramble siRNA PDGFRA MYC PDGFRA AURKA AURKA DIPG007siRNA2 PDGFRA NSC H3K27MsiRNA1and AURKA LAMTOR3 AURKA LAMTOR3 LAMTOR3 LIN28A LAMTOR3 LIN28A LIN28B LIN28A NSC-EV LIN28A MAP2K3 LIN28B LIN28B LIN28B MAP2K5 siRNA1 MAP2K3 MAP2K3 MAP3K2 MAP2K3 MAP3K7 MAP2K5 MAP2K5 MAPK7 MAP2K5 2 MAP3K2 siRNA1 siRNA2 MAP3K2 ZAK MAP3K7 MAP3K2 MAP3K7 MAPK7 MAP3K7 -

Insulin-Like Growth Factor Axis in Pregnancies Affected by Fetal Growth Disorders Aamod R
Nawathe et al. Clinical Epigenetics (2016) 8:11 DOI 10.1186/s13148-016-0178-5 RESEARCH Open Access Insulin-like growth factor axis in pregnancies affected by fetal growth disorders Aamod R. Nawathe1,2, Mark Christian3, Sung Hye Kim2, Mark Johnson1,2, Makrina D. Savvidou1,2 and Vasso Terzidou1,2* Abstract Background: Insulin-like growth factors 1 and 2 (IGF1 and IGF2) and their binding proteins (IGFBPs) are expressed in the placenta and known to regulate fetal growth. DNA methylation is an epigenetic mechanism which involves addition of methyl group to a cytosine base in the DNA forming a methylated cytosine-phosphate-guanine (CpG) dinucleotide which is known to silence gene expression. This silences gene expression, potentially altering the expression of IGFs and their binding proteins. This study investigates the relationship between DNA methylation of components of the IGF axis in the placenta and disorders in fetal growth. Placental samples were obtained from cord insertions immediately after delivery from appropriate, small (defined as birthweight <10th percentile for the gestation [SGA]) and macrosomic (defined as birthweight > the 90th percentile for the gestation [LGA]) neonates. Placental DNA methylation, mRNA expression and protein levels of components of the IGF axis were determined by pyrosequencing, rtPCR and Western blotting. Results: In the placenta from small for gestational age (SGA) neonates (n = 16), mRNA and protein levels of IGF1 were lower and of IGFBPs (1, 2, 3, 4 and 7) were higher (p < 0.05) compared to appropriately grown neonates (n =37).In contrast, in the placenta from large for gestational age (LGA) neonates (n = 20), mRNA and protein levels of IGF1 was not different and those of IGFBPs (1, 2, 3 and 4) were lower (p < 0.05) compared to appropriately grown neonates.