Peak Fitting Applied to Fourier Transform Infrared and Raman Spectroscopic Analysis of Proteins
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Future of Protein Secondary Structure Prediction Was Invented by Oleg Ptitsyn
biomolecules Review The Future of Protein Secondary Structure Prediction Was Invented by Oleg Ptitsyn 1, 1, 1 2 1,3 Daniel Rademaker y, Jarek van Dijk y, Willem Titulaer , Joanna Lange , Gert Vriend and Li Xue 1,* 1 Centre for Molecular and Biomolecular Informatics (CMBI), Radboudumc, 6525 GA Nijmegen, The Netherlands; [email protected] (D.R.); [email protected] (J.v.D.); [email protected] (W.T.); [email protected] (G.V.) 2 Bio-Prodict, 6511 AA Nijmegen, The Netherlands; [email protected] 3 Baco Institute of Protein Science (BIPS), Mindoro 5201, Philippines * Correspondence: [email protected] These authors contributed equally to this work. y Received: 15 May 2020; Accepted: 2 June 2020; Published: 16 June 2020 Abstract: When Oleg Ptitsyn and his group published the first secondary structure prediction for a protein sequence, they started a research field that is still active today. Oleg Ptitsyn combined fundamental rules of physics with human understanding of protein structures. Most followers in this field, however, use machine learning methods and aim at the highest (average) percentage correctly predicted residues in a set of proteins that were not used to train the prediction method. We show that one single method is unlikely to predict the secondary structure of all protein sequences, with the exception, perhaps, of future deep learning methods based on very large neural networks, and we suggest that some concepts pioneered by Oleg Ptitsyn and his group in the 70s of the previous century likely are today’s best way forward in the protein secondary structure prediction field. -

Chapter 4 the Three-Dimensional Structure of Proteins
Chapter 4 The Three-Dimensional Structure of Proteins Multiple Choice Questions 1. Answer: D All of the following are considered “weak” interactions in proteins, except: A) hydrogen bonds. B) hydrophobic interactions. C) ionic bonds. D) peptide bonds. E) van der Waals forces. 2. Answer: D In an aqueous solution, protein conformation is determined by two major factors. One is the formation of the maximum number of hydrogen bonds. The other is the: A) formation of the maximum number of hydrophilic interactions. B) maximization of ionic interactions. C) minimization of entropy by the formation of a water solvent shell around the protein. D) placement of hydrophobic amino acid residues within the interior of the protein. E) placement of polar amino acid residues around the exterior of the protein. 3. 3 Answer: A In the diagram below, the plane drawn behind the peptide bond indicates the: A) absence of rotation around the C—N bond because of its partial double-bond character. B) plane of rotation around the C—N bond. C) region of steric hindrance determined by the large C=O group. D) region of the peptide bond that contributes to a Ramachandran plot. E) theoretical space between –180 and +180 degrees that can be occupied by the and angles in the peptide bond. 4. Answer: D Which of the following best represents the backbone arrangement of two peptide bonds? A) C—N—C—C—C—N—C—C B) C—N—C—C—N—C C) C—N—C—C—C—N D) C—C—N—C—C—N Chapter 4 The Three-Dimensional Structure of Proteins E) C—C—C—N—C—C—C 5. -

Conformational Properties of Constrained Proline Analogues and Their Application in Nanobiology”
UNIVERSITAT POLITÈCNICA DE CATALUNYA DEPARTAMENT D’ENGINYERIA QUÍMICA “CONFORMATIONAL PROPERTIES OF CONSTRAINED PROLINE ANALOGUES AND THEIR APPLICATION IN NANOBIOLOGY” Alejandra Flores Ortega Supervisors: Dr. Carlos Alemán Llansó and Dr. Jordi Casanovas Salas. th Barcelona, 27 January 2009 “Chance is a word void of sense; nothing can exist without a cause”. François-Marie Arouet, Voltaire “Imagination will often carry us to worlds that never were. But without it, we go nowhere”. Carl Sagan iii ACKNOWLEDGEMENTS I would like to acknowledge to Dr. Carlos Aleman and Dr. Jordi Cassanovas Salas for an interesting research theme, and scientific support. I gratefully acknowledge to Dr. David Zanuy for interesting suggestions and strong discussions, without their support this would be an unfulfilled task. Also I, would like to address my thanks to all my colleagues in my group and department, specially Elaine Armelin for assiting me in many different ways. I thank not only my friends, but also colleagues for helping me to overcome the stressful time, without whom it would have been difficult to cope up. I wish to express my gratefulness to my parents, specially to my mother, María Esther, for all his care, and support. Also I will like to thanks to my friends and specially Jesus, Merches, Laura y Arturo. My PhD thesis have been finished for all this support. I am greatly indepted to Dr. Ruth Nussinov at NCI, Dr. Carlos Cativiela at the University of Zaragoza and Ana I. Jiménez at the “Instituto de Ciencias de Materiales de Aragon” for a collaborative effort. I wish to thank all my colleague in the “Chimie et Biochimie Théoriques, Faculté des Sciences et Techniques” in Nancy France, I will be grateful to have worked with : Pr. -

2. Proteins Have Hierarchies of Structure
27 2. Proteins have Hierarchies of Structure Protein structure is usually described at four different levels (Fig. II.2.1). The first level, called the primary structure, describes the linear sequence of the amino acids in the chain. The different primary structures correspond to the different sequences in which the amino acids are covalently linked together. The secondary structure describes two common patterns of structural repetition in proteins: the coiling up into helices of segments of the chain, and the pairing together of strands of the chain into β-sheets. The tertiary structure is the next higher level of organization, the overall arrangement of secondary structural elements. The quaternary structure describes how different polypeptide chains are assembled into complexes. Figure II.2.1. Different levels of protein structure. A protein chain’s primary structure is its amino acid sequence. Secondary structure consists of the regular organization of helices and sheets. The example shown is a schematic representation of an α helix. Tertiary structure is a polypeptide chain’s three- dimensional native conformation, which often involves compact packing of secondary structure elements. The example given in the figure is a schematic drawing of one of the four polypeptide chains (subunits) of hemoglobin, the protein that transports oxygen in the blood. α-helices along the chain are represented as cylinders. N is the amino terminus and C is the carboxyl terminus of the polypeptide chain. Quaternary structure is the arrangement of multiple polypeptide chains (subunits) to form a functional biomolecular structure. The figure shows the quaternary arrangement of four subunits to form the functional hemoglobin molecule. -

Introduction to Proteins and Amino Acids Introduction
Introduction to Proteins and Amino Acids Introduction • Twenty percent of the human body is made up of proteins. Proteins are the large, complex molecules that are critical for normal functioning of cells. • They are essential for the structure, function, and regulation of the body’s tissues and organs. • Proteins are made up of smaller units called amino acids, which are building blocks of proteins. They are attached to one another by peptide bonds forming a long chain of proteins. Amino acid structure and its classification • An amino acid contains both a carboxylic group and an amino group. Amino acids that have an amino group bonded directly to the alpha-carbon are referred to as alpha amino acids. • Every alpha amino acid has a carbon atom, called an alpha carbon, Cα ; bonded to a carboxylic acid, –COOH group; an amino, –NH2 group; a hydrogen atom; and an R group that is unique for every amino acid. Classification of amino acids • There are 20 amino acids. Based on the nature of their ‘R’ group, they are classified based on their polarity as: Classification based on essentiality: Essential amino acids are the amino acids which you need through your diet because your body cannot make them. Whereas non essential amino acids are the amino acids which are not an essential part of your diet because they can be synthesized by your body. Essential Non essential Histidine Alanine Isoleucine Arginine Leucine Aspargine Methionine Aspartate Phenyl alanine Cystine Threonine Glutamic acid Tryptophan Glycine Valine Ornithine Proline Serine Tyrosine Peptide bonds • Amino acids are linked together by ‘amide groups’ called peptide bonds. -

Detection of Trans-Cis Flips and Peptide-Plane Flips in Protein Structures
research papers Detection of trans–cis flips and peptide-plane flips in protein structures ISSN 1399-0047 Wouter G. Touw,a* Robbie P. Joostenb and Gert Vrienda* aCentre for Molecular and Biomolecular Informatics, Radboud University Medical Center, Geert Grooteplein-Zuid 26-28, 6525 GA Nijmegen, The Netherlands, and bDepartment of Biochemistry, Netherlands Cancer Institute, Plesmanlaan 121, 1066 CX Amsterdam, The Netherlands. *Correspondence e-mail: [email protected], Received 4 February 2015 [email protected] Accepted 27 April 2015 A coordinate-based method is presented to detect peptide bonds that need Edited by G. J. Kleywegt, EMBL–EBI, Hinxton, correction either by a peptide-plane flip or by a trans–cis inversion of the peptide England bond. When applied to the whole Protein Data Bank, the method predicts 4617 trans–cis flips and many thousands of hitherto unknown peptide-plane flips. Keywords: peptide conformation; cis peptide A few examples are highlighted for which a correction of the peptide-plane bond; structure validation; structure correction. geometry leads to a correction of the understanding of the structure–function relation. All data, including 1088 manually validated cases, are freely available and the method is available from a web server, a web-service interface and through WHAT_CHECK. 1. Introduction Peptide bonds connect adjacent amino acids in proteins. The partial double-bond character of the peptide bond restricts its torsion. The dihedral angle ! (CiÀ1—CiÀ1—Ni—Ci ) typically has values around 180 (trans)or0 (cis), although exceptions are possible (Berkholz et al., 2012). The Ci–1—Ci distance is around 3.81 A˚ in the trans conformation and around 2.94 A˚ in the cis conformation. -
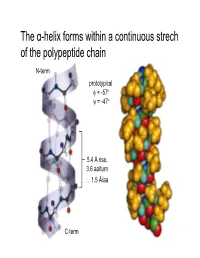
The Α-Helix Forms Within a Continuous Strech of the Polypeptide Chain
The α-helix forms within a continuous strech of the polypeptide chain N-term prototypical φ = -57 ° ψ = -47 ° 5.4 Å rise, 3.6 aa/turn ∴ 1.5 Å/aa C-term α-Helices have a dipole moment, due to unbonded and aligned N-H and C=O groups β-Sheets contain extended (β-strand) segments from separate regions of a protein prototypical φ = -139 °, ψ = +135 ° prototypical φ = -119 °, ψ = +113 ° (6.5Å repeat length in parallel sheet) Antiparallel β-sheets may be formed by closer regions of sequence than parallel Beta turn Figure 6-13 The stability of helices and sheets depends on their sequence of amino acids • Intrinsic propensity of an amino acid to adopt a helical or extended (strand) conformation The stability of helices and sheets depends on their sequence of amino acids • Intrinsic propensity of an amino acid to adopt a helical or extended (strand) conformation The stability of helices and sheets depends on their sequence of amino acids • Intrinsic propensity of an amino acid to adopt a helical or extended (strand) conformation • Interactions between adjacent R-groups – Ionic attraction or repulsion – Steric hindrance of adjacent bulky groups Helix wheel The stability of helices and sheets depends on their sequence of amino acids • Intrinsic propensity of an amino acid to adopt a helical or extended (strand) conformation • Interactions between adjacent R-groups – Ionic attraction or repulsion – Steric hindrance of adjacent bulky groups • Occurrence of proline and glycine • Interactions between ends of helix and aa R-groups His Glu N-term C-term -

Mitigation of Terrestrial Radar Interference in L-Band Spaceborne Microwave Radiometers
Mitigation of Terrestrial Radar Interference in L-Band Spaceborne Microwave Radiometers J.R. Piepmeier and F.A. Pellerano Microwave Instrument Technology Branch NASA's Goddard Space Flight Center Greenbelt, MD 2 1 146 Email: [email protected] Abstract-Terrestrial radars operating in the 1215-1400 MHz Hydros missions [lo]. Given the impacts, three aspects to radio-location and navigation spectrum allocation are important mitigation are proposed: survivability, operability, and excis- for air traffic safety, homeland security, and national defense. For ability; or SOE. Survivability deals with avoiding damage low-frequency observations of soil moisture and ocean salinity, Earth-observing microwave radiometers are allocated Earth- from RFI. This means proper filtering and limiting. A ra- Exploration Satellite Service (EESS) spectrum for operating at diometer achieves operability when it can measure, without 1400-1427 MHz. The proximity of powerful long-range radars to error, the antenna or brightness temperature in the presence of the passive allocation makes observing a challenge. Three aspects interference. Selective receivers operating in quite spectrum of mitigation to RFI are discussed in this paper: survivability, are required for operability. When filtering and frequency operability, and excisability (SOE). Modeling and simulations of NASA's Hydros and Aquarius radiometers were performed selection is not enough to avoid interference, then the RFI to examine the impacts of radar interference. The results are might be excisable. A number of techniques have been pro- applied to the three aspects of mitigation SOE and the affects posed and demonstrated recently [Ill-[14]. These fall into on the radiometer requirements are discussed. three basic categories: temporal, spectral, and statistical - some techniques are a combination of two or more. -

Terahertz Induced Non-Linear Electron Dynamics in Nanoantenna Coated Semiconductors at the Sub-Picosecond Timescale
AN ABSTRACT OF THE DISSERTATION OF Andrew Stickel for the degree of Doctor of Philosophy in Physics presented on August 10, 2016. Title: Terahertz Induced Non-linear Electron Dynamics in Nanoantenna Coated Semiconductors at the Sub-picosecond Timescale Abstract approved: Yun-Shik Lee This dissertation is an exploration of the material response to Terahertz (THz) radiation. Specifically we will explore the ultrafast electron dynamics in the non- perturbative regime in semiconductors that have been patterned with nanoantenna arrays using broadband, high intensity, THz radiation. Three main semiconductor materials will be studied in this work. The first is VO2 which undergoes a phase transition from an insulator, when it is below 67◦ C, to a metal, when it is above 67◦ C. The second and third materials are Si and GaAs which are two of the most commonly used semiconductors. We study the insulator to metal transition (IMT) of VO2 and its response to high field THz radiation. The near room temperature IMT for VO2 makes it a very promising material for electrical and photonic applications. We demonstrate that with high field THz the IMT transition can be triggered. This transition is induced on a sub-cycle timescale. We also demonstrate a THz field dependent reduction in the transition temperature for the IMT when transitioning from both below Tc to above as well as from above Tc to below. This transition is not equal for the above and below cases and leads to a narrowing of the hysteresis curve of the IMT. The thin film Fresnel coefficients, along with a phenomenological model developed for the nanoantenna patterned VO2, are also used to calculate the sheet conductivity of the VO2 sample. -

And Beta-Helical Protein Motifs
Soft Matter Mechanical Unfolding of Alpha- and Beta-helical Protein Motifs Journal: Soft Matter Manuscript ID SM-ART-10-2018-002046.R1 Article Type: Paper Date Submitted by the 28-Nov-2018 Author: Complete List of Authors: DeBenedictis, Elizabeth; Northwestern University Keten, Sinan; Northwestern University, Mechanical Engineering Page 1 of 10 Please doSoft not Matter adjust margins Soft Matter ARTICLE Mechanical Unfolding of Alpha- and Beta-helical Protein Motifs E. P. DeBenedictis and S. Keten* Received 24th September 2018, Alpha helices and beta sheets are the two most common secondary structure motifs in proteins. Beta-helical structures Accepted 00th January 20xx merge features of the two motifs, containing two or three beta-sheet faces connected by loops or turns in a single protein. Beta-helical structures form the basis of proteins with diverse mechanical functions such as bacterial adhesins, phage cell- DOI: 10.1039/x0xx00000x puncture devices, antifreeze proteins, and extracellular matrices. Alpha helices are commonly found in cellular and extracellular matrix components, whereas beta-helices such as curli fibrils are more common as bacterial and biofilm matrix www.rsc.org/ components. It is currently not known whether it may be advantageous to use one helical motif over the other for different structural and mechanical functions. To better understand the mechanical implications of using different helix motifs in networks, here we use Steered Molecular Dynamics (SMD) simulations to mechanically unfold multiple alpha- and beta- helical proteins at constant velocity at the single molecule scale. We focus on the energy dissipated during unfolding as a means of comparison between proteins and work normalized by protein characteristics (initial and final length, # H-bonds, # residues, etc.). -

PA28: New Insights on an Ancient Proteasome Activator
biomolecules Review PA28γ: New Insights on an Ancient Proteasome Activator Paolo Cascio Department of Veterinary Sciences, University of Turin, Largo P. Braccini 2, 10095 Grugliasco, Italy; [email protected] Abstract: PA28 (also known as 11S, REG or PSME) is a family of proteasome regulators whose members are widely present in many of the eukaryotic supergroups. In jawed vertebrates they are represented by three paralogs, PA28α, PA28β, and PA28γ, which assemble as heptameric hetero (PA28αβ) or homo (PA28γ) rings on one or both extremities of the 20S proteasome cylindrical structure. While they share high sequence and structural similarities, the three isoforms significantly differ in terms of their biochemical and biological properties. In fact, PA28α and PA28β seem to have appeared more recently and to have evolved very rapidly to perform new functions that are specifically aimed at optimizing the process of MHC class I antigen presentation. In line with this, PA28αβ favors release of peptide products by proteasomes and is particularly suited to support adaptive immune responses without, however, affecting hydrolysis rates of protein substrates. On the contrary, PA28γ seems to be a slow-evolving gene that is most similar to the common ancestor of the PA28 activators family, and very likely retains its original functions. Notably, PA28γ has a prevalent nuclear localization and is involved in the regulation of several essential cellular processes including cell growth and proliferation, apoptosis, chromatin structure and organization, and response to DNA damage. In striking contrast with the activity of PA28αβ, most of these diverse biological functions of PA28γ seem to depend on its ability to markedly enhance degradation rates of regulatory protein by 20S proteasome. -

Disarmament Conference'
University of Nebraska at Omaha DigitalCommons@UNO Kabul Times Digitized Newspaper Archives 7-28-1965 Kabul Times (July 28, 1965, vol. 4, no. 104) Bakhtar News Agency Follow this and additional works at: https://digitalcommons.unomaha.edu/kabultimes Part of the International and Area Studies Commons Recommended Citation Bakhtar News Agency, "Kabul Times (July 28, 1965, vol. 4, no. 104)" (1965). Kabul Times. 1025. https://digitalcommons.unomaha.edu/kabultimes/1025 This Newspaper is brought to you for free and open access by the Digitized Newspaper Archives at DigitalCommons@UNO. It has been accepted for inclusion in Kabul Times by an authorized administrator of DigitalCommons@UNO. For more information, please contact [email protected]. ~!'. - _. .,.. -::'~ _:.~---- ';'- ' . _.: . ---~ _._ ....... :$2:_ 3 ;c: ... ce,,~ -. : ': ': . :; ',-'•.f - . -. ..:. " -' , -.'. -. .: -~. - . - . .. ,: - '. ••.t • - -.:- --.:'~ '~-' ...- '. -. .. -. ." ..- - -- • : : '- . " . .. --- '" .. , , : - - . -... _. :-. -: :-- - . '. ., . -. ~ - ' . .. - . '. , . , · ., .' .- ---. ~ . ,; __ 7-_ ..- • -. - .. - • < ,.., ,- .. :. '.' , . if '. NEWS STALLS . -.... -, . ~ -"- - ~. THE WEATHER "-". .., . KABUL TIMES JULY 27, 1965 Yesterday'S Tempera&are . ~, < • , .'t,. Max, + 31'C, Minimum 12°C· I·,' .: ..~ , Sun sets· today at 6:54 P.DL Belgian En-voy He.r-e.NamedI '- , ,Nasser De'njes VARAIr Force· suD. rjses tomorrow at 5:03 am· - Tomorrow's Outlook: Cloudy ~ -.. KA~UL, 27.~The .' ".' ',' 'Tr'ied To'Bomb His~Re$idence' July Ministry . of 'Foreign Affairs smd :yesterday j VOL. rv,NO. 104 KABUL'WEDNESDAY,. .JULY ·28"(ASA'D:··6~1344;··S.H.) :: .,:, -:'.:'. , . =;:.0 .....- . :"'-, '. '.;-- / . s . .(DPA):~ ~~~~~~_~~~~~ ,_~ ~".~~~.~~~c~,~~:~._.~~.~.~:~~~~~~~~~~~~.~.~_~.~.,~:=.~~.C=E~.A~.!~:! ~.:~~~ '·:.ALExANDRIA, JulY' ·27) that Jean Leroy, .the Belgian Am e' . , .. , .. .. pRESIDENT Gamal Abci~l.Nasser.1?f.t~e uAR has ~tegori. bassado!' in N~w DeIhi, -has been '. , cally d~n1ed th!lt an attelJi:)t~~ been made by aU' force, appointed to act siniultaneously as ·1Ii......· ,.