Palaeontological Society of Japan
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Traditional Crafts of Kumamoto Various Traditional Crafts Are Used in Everyday Life in Kumamoto
Traditional Crafts of Kumamoto Various traditional crafts are used in everyday life in Kumamoto. These crafts are born from Kumamoto’s natural environment, the skills Traditional Crafts of Kumamoto of craftsmen, and the ingenuity used by locals in their daily lives. Kumamoto’s handicrafts are created through communication between Craft items that originate from Kumamoto and were handed down for the craft creators and the craft users. They are found in a variety of generations are designated “Traditional Crafts of Kumamoto.” To receive this places and used in a variety of ways. designated, the craft must be made using traditional techniques and must have over 30 years of history. There are about 90 such designated crafts in Kumamoto, including metalwork, ceramics, woodwork, bamboo crafts, dying and weaving, paper products, and traditional toys. Japan’s Nationally Designated Crafts To be deemed a “Nationally Designated Craft,” the traditional skills or techniques used to make the craft must have over 100 years of history, and must have developed in a fixed region with more than 10 organizations or 30 individual craftsmen currently engaged in the production of the craft. Over 200 crafts in Japan have been declared Nationally Designated Crafts, including Kyo and Arita ware pottery, and Wajima-style lacquerware. In Kumamoto, Shodai pottery, Amakusa ceramics, and Higo inlay metalwork all received this distinction in March 2003. In December 2013, Yamaga lanterns were the fourth craft from Kumamoto to be designated. 1 Higo-Zogan Metalwork Metalwork in Kumamoto includes the following crafts: Higo-zogan, which originated from sword accessories; Kawashiri and Hitoyoshi-Kuma cutting tools, such as kitchen knives, farm hoes and sickles; and swords, the production of which dates back 750 years ago to the Kamakura Period. -

Storm Surge Damage Caused by Typhoon No. 9918 in the Area of the Shiranui Sea
DISASTER PREVENSION MEASURES Storm Surge Damage caused by Typhoon No. 9918 in the Area of the Shiranui Sea Lessons learned Kiyoshi TAKIKAWA Member of JSCE, Dr. of Eng., Professor, Research and Education Center of Coastal Environmental Science, Kumamoto University Damage overview was estimated at ¥100 billion, the worst ever in the pre- fecture. Typhoon No. 18 of 1999 (Typhoon 9918) cut across Immediately after the event, Kumamoto University the Kyushu and Chugoku regions on September 24, began a site survey, joining with the JSCE's coastal engi- leaving widespread damage in its wake. The maximum neering research group to inspect the damage in detail. instantaneous wind speed reached a record high of 66.2 Information about the damage was collected on the fol- meters per second at the Ushibuka Weather Station run lowing day, September 25. by Kumamoto Meteorological Observatory. In The results of the survey were made public at the Kumamoto Prefecture, the Matsuai District of Shiranui Coastal Engineering Symposium, held in Yonago City Town lies at the innermost extremity of an inlet in on November 15, 1999. On the day preceding the sym- Shiranui (Yatsushiro) Bay. Here, storm-driven tides posium, the JSCE's Coastal Engineering Committee held flooded over low-lying land and claimed 12 lives. These an emergency briefing and panel discussion, at which were the first victims of a storm surge in the country lively discussion of the academic challenges arising since the Ise Bay Typhoon of September 1959, so the from studies of the damage and the technical focus of loss of life was a shock to us all. -

By Municipality) (As of March 31, 2020)
The fiber optic broadband service coverage rate in Japan as of March 2020 (by municipality) (As of March 31, 2020) Municipal Coverage rate of fiber optic Prefecture Municipality broadband service code for households (%) 11011 Hokkaido Chuo Ward, Sapporo City 100.00 11029 Hokkaido Kita Ward, Sapporo City 100.00 11037 Hokkaido Higashi Ward, Sapporo City 100.00 11045 Hokkaido Shiraishi Ward, Sapporo City 100.00 11053 Hokkaido Toyohira Ward, Sapporo City 100.00 11061 Hokkaido Minami Ward, Sapporo City 99.94 11070 Hokkaido Nishi Ward, Sapporo City 100.00 11088 Hokkaido Atsubetsu Ward, Sapporo City 100.00 11096 Hokkaido Teine Ward, Sapporo City 100.00 11100 Hokkaido Kiyota Ward, Sapporo City 100.00 12025 Hokkaido Hakodate City 99.62 12033 Hokkaido Otaru City 100.00 12041 Hokkaido Asahikawa City 99.96 12050 Hokkaido Muroran City 100.00 12068 Hokkaido Kushiro City 99.31 12076 Hokkaido Obihiro City 99.47 12084 Hokkaido Kitami City 98.84 12092 Hokkaido Yubari City 90.24 12106 Hokkaido Iwamizawa City 93.24 12114 Hokkaido Abashiri City 97.29 12122 Hokkaido Rumoi City 97.57 12131 Hokkaido Tomakomai City 100.00 12149 Hokkaido Wakkanai City 99.99 12157 Hokkaido Bibai City 97.86 12165 Hokkaido Ashibetsu City 91.41 12173 Hokkaido Ebetsu City 100.00 12181 Hokkaido Akabira City 97.97 12190 Hokkaido Monbetsu City 94.60 12203 Hokkaido Shibetsu City 90.22 12211 Hokkaido Nayoro City 95.76 12220 Hokkaido Mikasa City 97.08 12238 Hokkaido Nemuro City 100.00 12246 Hokkaido Chitose City 99.32 12254 Hokkaido Takikawa City 100.00 12262 Hokkaido Sunagawa City 99.13 -
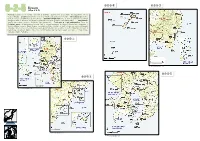
6-2-5-④ 6-2-5-③ Kyusyu 6-2-5 (Map 6-2-5) Province: Kumamoto Pref
6-2-5-④ 6-2-5-③ Kyusyu 6-2-5 (Map 6-2-5) Province: Kumamoto Pref. at west, Oita Pref. at northeast, Miyazaki Pref. at southeast, and Kagoshima Pref. at south of Kyushu Location: Kyushu lies at west of Shikoku and southwest of Honshu Air temperature: 17.8˚C (annual average, at Ushibuka City, Kumamoto.) Seawater temperature: 22.9˚C, 22.2 ˚C and 20.9 ˚C (annual average, at east off Aburatsu, southwest off Kushikijima (Is.) and Yatsushiro Sea, respectively) Precipitation: 2,027.9 mm (annual average, Ushibuka City, Kumamoto) Total area of coral communities: 581.8 ha Protected areas: Unzen-Amakusa National Park: at around Amakusa, including 3 Marine Park Zones and 2 Protected Water Surfaces; Nippo Kaigan Quasi-National Park: coastline at south of Oita and north of Miyazaki, including 2 Marine Park Zones; Nichinan Kaigan Quasi-National Park: coastline at south of Miyazaki and east of Kagoshima, including 1 Marine Park Zone; Kirishima-Yaku National Park: a part of coastline in Kagoshima, including 2 Marine Park Zones. 6-2-5-① *“号”on this map means“site”. 6-2-5-⑤ 6-2-5-② *“号”on this map means“site”. 6-2-5-② 6-2-5-① *“号”on this map means“site”. 6-2-5-④ 6-2-5-③ *“号”on this map means“site”. 6-2-5-⑤ 06 Coral Reefs of Japan a. Kumamoto Prefecture (Map 6-2-5-①) Satoshi Nojima 1 Corals and coral reefs Photo. 1. Tabulate Acropora dominant community in Kuwashima 1. Geographical features (Is.), Ushibuka City, Kumamoto Prefecture. On the west of Kyushu in Kumamoto Prefecture lie the Amakusa Islands. -

Palaeontological Society of Japan
Transactions and Proceedings of the Palaeontological Society of Japan New Series No. 88 Palaeontological Society of Japan December 20, 1972 ---------------------·-· Editor: Takashi HAMADA Associate Editor: Yasuhide IWASAKI Officers for 1971 - 1972 President : Tokio SHIKAMA Councillors(* Executives): Kiyoshi ASAND*, Kiyotaka CHINZEI*, Takashi HAMADA*, Tetsuro HANAI*, Kotara HAT AI, ltaru HAY AMI, Koichiro lCHIKA WA, Taro KANAYA, Kametoshi KANMERA, Tamio KOTAKA, Tatsuro MATSUMOTO*, Hiroshi OZAKI*, Tokio SHIKAMA*, Fuyuji TAKA!*, Yokichi TAKAYANAGI Secretaries: Wataru HASHIMOTO, Saburo KANNO Executive Committee General Affairs: Tetsuro HANAI, Naoaki AOKI Membership : Kiyotaka CHINZEI, Toshio KOIKE Finance: Fuyuji T AKAI, Hisayoshi lGO Planning: Hiroshi OZAKI, Kazuo AsAMA Publications Transactions: Takashi HAMADA, Yasuhide IWASAKI Special Papers: Tatsuro MATSUMOTO, Tomowo OZAWA '' Fossils" : Kiyoshi AsANO, Toshiaki TAKAYAMA All communications relating to this journal should be addressed to the PALAEONTOLOGICAL SOCIETY OF JAPAN c/o Business Center for Academic Societies, Japan Yayoi 2-4-16, Bunkyo-ku, Tokyo 113, Japan. Sole agent: University of Tokyo Press, Hongo, Tokyo Trans. Proc. Palaecint. Soc. Japan, N. S., No. 88, pp. 447-457, pl. 54, December 20, 1972 607. UPPER TRIASSIC CEPHALOPODS FROM THE TANOURA DISTRICT, KUMAMOTO PREFECTURE, JAPAN* T AKESHI ISHIBASHI Department of Geology, Faculty of Science, Kyushu University Jm;fs:~llliinit!!JrRO)J:ll!I.:::.~*~~.:Em = lm**~HI\. mim!i!Wll::~:tn-t7.> .=:.t~l±liilll! :t,;.tCJ'!liili"'JI.t IJ imllit...t-:. \!J:.lEID~#it-tt..~ifi*· ;;t-l,~W nt. ~:Pii stm~~ll'.t.. t-:., ? ~~1i 1 f!H1¥rf<1l't'<l:>oo .:.tt,~O)iji.lEmri'l'ft~'t'li<I:>.Oil:, 8*0).:::.:\ilf..t 1HJJII.> "Cffl'.!i' <Ht.7.> ~lim t <!:> 1J, i!ti.lEfflitJ#JtlfO)lVf~i;:l;tiJtli:"t' <1; .o. -

Distribution of Sediment, Benthic Foraminifera and Mercury in the South Yatsushiro Sea, Kyushu, Japan
Jurnal of Coastal Development ISSN : 1720-1041 Volume 11 Number 3, June 2008 : 104-112 DISTRIBUTION OF SEDIMENT, BENTHIC FORAMINIFERA AND MERCURY IN THE SOUTH YATSUSHIRO SEA, KYUSHU, JAPAN Rifardi Department. of Marine Science, Faculty of Fisheries and Marine Sciences, Riau University Indonesia. Kampus Bina Widya Sp. Panam Pekanbaru-Riau Indonesia Received :April, 23, 2008 ; Accepted: May, 29, 2008 ABSTACTS Bottom sediment samples were collected at 74 stations in the South Yatsushiro Sea, off the west coast of central Kyushu Island, Japan using gravity corer. 62 core samples were used for bottom sediments analysis and for determination of mercury content. Among these samples, only 5 core samples were used for this study in an attempt to clarify the relationship between the vertical distributions of benthic foraminiferal assemblages (Bulimina denudata) and mercury contents in core sediments. The distribution pattern of sediment median diameter and of the maximum mercury content at some layers of each core show that the fine-grained sediment polluted by mercury were transported both northeastward and southward by weak longshore currents and spread toward north and west across the northern and souhtern part of the South Yatsushiro Sea. Bulimina denudata shows the highest frequency at every layer which has high mercury content ranging from 0.14 to 3.46 ppm, and is comparatively low at all unpolluted layers. The frequency of Bulimina denudata slightly decreased at the layers showing more than 3 ppm. All these signals suggest that Bulimina denudata does not prefer the bottom sediments showing high mercury contents but tolerates mercury pollution. Keywords: Bottom sediment, benthich forminifera, mercury. -

A Prosperous Future Starts Here
A prosperous future starts here 100% of this paper was made using recycled paper 2018.4 (involved in railway construction) Table of Lines Constructed by the JRTT Contents Tsukuba Tokyo Area Lines Constructed by JRTT… ……………………… 2 Sassho Line Tsukuba Express Line Asahikawa Uchijuku JRTT Main Railway Construction Projects……4 Musashi-Ranzan Signal Station Saitama Railway Line Maruyama Hokkaido Shinkansen Saitama New Urban Musashino Line Tobu Tojo Line Urawa-Misono Kita-Koshigaya (between Shin-Hakodate-Hokuto Transit Ina Line Omiya Nemuro Line Shinrin-Koen and Sapporo) ■ Comprehensive Technical Capacity for Railway Sapporo Construction/Research and Plans for Railway Tobu Isesaki Line Narita SKY ACCESS Line Construction… ………………………………………………6 Hatogaya (Narita Rapid Rail Acess Line) Shiki Shin-Matsudo Hokuso Railway Hokuso Line ■ Railway Construction Process… …………………………7 Takenotsuka Tobu Tojo Line Shin-Kamagaya Komuro Shin-Hakodatehokuto Seibu Wako-shi Akabane Ikebukuro Line Imba Nihon-Idai Sekisho Line Higashi-Matsudo Narita Airport Hakodate …… Kotake-Mukaihara Toyo Rapid Construction of Projected Shinkansen Lines 8 Shakujii-Koen Keisei-Takasago Hokkaido Shinkansen Aoto Nerima- Railway Line Nerima Takanodai Ikebukuro Keisei Main Line (between Shin-Aomori and Shin-Hakodate-Hokuto) Hikifune Toyo- Tsugaru-Kaikyo Line Seibu Yurakucho Line Tobu Katsutadai ■ Kyushu Shinkansen… ………………………………………9 Tachikawa Oshiage Ueno Isesaki Line Keio Line Akihabara Nishi-Funabashi Shinjuku … ………………………………… Odakyu Odawara Line Sasazuka ■ Hokuriku Shinkansen 10 Yoyogi-Uehara -

Ama Kusa Tamana / Yamaga / Kikuchi Kumamoto Central Region Aso Kuma Yatsushiro / Minamata
Hakata Fukuoka Airport Kokura Nagasaki Expressway Tsuetate Yufuin TAMANA / YAMAGA / KIKUCHI Kurokawa Nankan IC Kikuchi Castle Ruins Nichirinji Temple Aso/Kuju National Park Yamaga Ikeyama Fountainhead Greenland Kikuchi Kikusui IC Yachiyoza ▲ Daikanbou Nakae Kagura (Sacred music and dancing) Shin-Tamana Station Kikuchi Gorge Nagasu Tamana Ueki IC Uchinomaki Komezuka Kyushu Expressway Cuddly Dominion Taketa Ueki Aso Shrine Yoh Shomei Picture Book Museum ▲Mt. Aso (Crater) Kumamoto Castle Aso Farmland Taira Reigando Cave Kusasenri Shimada Art Museum Ozu Monkey Show Theatre Minami-Aso Spa District Shimabara Peninsula Ariake Sea (Tochinoki, Jigoku, Tarutama, Yunotani) Yurugiga-Ike Minami-Aso Railway Shirakawa Fountainhead Prefectural Traditional Airport Aso Kumamoto Shimabara Crafts Center Kumamoto IC Nagasaki Kumamoto Station Mashiki-Kumamoto Airport IC ASO Mogi Uto Mifune IC Takachiho Todoroki Suizenji Jojuen Park Yamato AMA Kuchinotsu Misumi Fountainhead Seiwa Bunrakukan Hall Mamihara Matsubase IC Tsujunkyo Bridge KUSA Reitaikyo Bridge Dolphin Watching Oyano Yatsushiro Miyahara KUMAMOTO CENTRAL REGION Tomioka Oniike Castle Ruins Shin-Yatsushiro Station Sendan-Todoro Suspension Bridge Amakusa Airport Matsushima Shohinken Yatsushiro Station Yatsushiro IC Shiiba Yatsushiro-Minami IC Hondo Gokanosho Amakusa Kamishima Myoken Ura Shiranui Sea Shimoda Hinagu-Onsen Station Shrine Yatsushiro Amakusa Shimoshima Odo Hinagu IC Itsuki / Gokanosho Prefectural Natural Park Oe Catholic Cathedral Sakitsu Tanoura IC Itsuki Catholic Cathedral Nakada Ashikita IC Hinagu Utase-Bune Kyusendo Cave Yunomae Yuyama Kuma River Hitoyoshi IC Goshoura Rapid Shooting Hitoyoshi Castle Ruins Ichifusa Dam Ushibuka HIsatsu Orange Railway Kumagawa Railway Hitoyoshi Yunoko Shin-Minamata Station Minamata Station Kuranomoto Yunotsuru Minamata KUMA YATSUSHIRO / Miyazaki Kagoshima MINAMATA Ebino IC Legend National Highway Expressway JR Sea Route Aviation Shinkansen How to use the Map Fukuoka Beijing Locations are indicated next to each name. -

Notes on the Distribution of the Vandenboschia Radicans Complex (Hymenophyllaceae) in Japan
Bull. Natl. Mus. Nat. Sci., Ser. B, 35(2), pp. 71–89, June 22, 2009 Notes on the Distribution of the Vandenboschia radicans Complex (Hymenophyllaceae) in Japan Atsushi Ebihara Department of Botany, National Museum of Nature and Science, Amakubo 4–1–1, Tsukuba, 305–0005 Japan E-mail: [email protected] Abstract The distribution of eight species and four hybrids included in the Vandenboschia radi- cans complex within Japan is reviewed. Distribution maps for each taxon are provided based on herbarium specimens, which are listed in order of municipality code. Key words:Distribution, Hymenophyllaceae, Japan, Vandenboschia. Introduction were embedded in V. ϫstenosiphon and V. ϫ Vandenboschia subgen. Vandenboschia (Hy- quelpaertensis, respectively because it is diffi- menophyllaceae) distributed in world temperate/ cult to discriminate amphidiploids from sterile subtropical regions encompasses a number of hybrids sharing the same genome combinations morphologically ill-defined species, and they are (but different genome dosage). As shown in the tentatively lumped under a species complex discriminant analysis (Ebihara et al., 2009), some named “Vandenboschia radicans complex”. As of the redefined taxa morphologically overlap for Japanese members of the complex, Ebihara with each other. Therefore, descriptions and keys et al. (2005a) clarified their biological entities by do not always provide sufficient information for DNA/ploidy analyses, and Ebihara et al. (2009) their identification. Digital images of examined reexamined their taxonomic properties. Since the specimens (only those deposited in TNS), to number of samples analyzed in these studies is compensate the difficulty, are accessible via on- still limited, it is necessary to examine as many line database (http://www.vspecimens.net/). -

Responses of GSI to the 2016 Kumamoto Earthquake Disaster Management Office, Planning Department
Responses of GSI to the 2016 Kumamoto Earthquake 15 Responses of GSI to the 2016 Kumamoto Earthquake Disaster Management Office, Planning Department (Published online: 28 December 2016) Abstract Geospatial Information Authority of Japan (GSI) provides geospatial information to grasp the disaster situation to relevant organizations such as ministries and government offices, local public organizations and the like to help contribute in the lifesaving and rescue activities as well as recovery and reconstruction efforts when a large-scale natural disaster occurs. GSI provided geospatial information to concerned administrative authorities including the Ministry of Land, Infrastructure, Transport and Tourism (MLIT), local government and related organ, in connection with the 2016 Kumamoto earthquake. This paper includes an overview of the disaster and reports on GSI’s main responses. 1. Overview of the Kumamoto Earthquake The 2016 Kumamoto earthquake (hereinafter referred to as “the Kumamoto earthquake”) included a magnitude 6.5 earthquake which occurred on April 14 at 21:26 JST, followed by a magnitude 7.3 earthquake which occurred on April 16 at 1:25 JST. Thus, these events produced a maximum seismic intensity 7 in Kumamoto. After April 14, 21:26 JST, 2 earthquakes with a maximum seismic intensity of a little over 6 and 3 earthquakes with a maximum seismic intensity of a little less than 6 occurred. Damages caused by the Kumamoto earthquake Fig. 1 Seismic intensity map (April 14) include: 98 human casualties, 830 seriously injured, 1,491 (Japan Meteorological Agency, 2016) people who suffered minor injuries, 8,198 buildings were completely destroyed, 29,761 were partially destroyed and 138,102 suffered minor damage. -

Amakusa Takarajima Tourist Association 出水駅~阿久根へ 15-7 Chuo-Shinmachi, Amakusa City, Kumamoto Pref
Mt.Unzen to Kumamoto Shimabara Okoshiki Coast Nature Park City Rest Stop ‘Uto Marina’ Shimabara Bay sa M maku isum Uto R A i Lin World Heritage J e 宇土市 Shimabara Peninsula Uto City to Kumamoto Amakusa Shiro Tokisada Giant According to oral tradition, the Amakusa-Shimabara Rebellion Amakusa Shiro (1637-1638) was said to be an uprising of Christians, but Statue Nishi-Arie Misumi Nishiko Historic Port Mt. Otake Todoroki Spring actually it was a peasant riot not directly motivated by religious ▲ causes. It happened because the local peasants were burdened An island located between Amakusa Nagasaki Prefecture and Shimabara, where Christian with unreasonably heavy taxes under the Tokugawa Shogunate. insurgents from both sides Matsubase met to discuss strategy for Matsubase Even after the rebellion was crushed, Shigenari Suzuki, an World Heritage IC the Amakusa-Shimabara Rest Stop ‘Sun Pearl’ 宇城市 Town official in charge of the Amakusa Islands, is said to have asked Ruins of Rebellion. Hara Castle ▲ Uki City the shogun to ease the tax burden on his people, explaining Mt. Misumidake Rest Stop ‘Shiranui Spa’ ▲ that the requisite amount of rice could not be grown in Mt. Shibaoyama Amakusa’s infertile soils. Kuchinotsu Port Misumi Yushima Island Misumi Branch Office,Uki City Office ( ‘Collusion’ Island) Tobase Island Shimatetsu Nagao Shrine to Mogi(Nagasaki) Ferry e n Ogawa i ン Spa Thalasso L Fast Ship Ariake Octopus e イ Town Wakamiya Beach , Amakusa Shiro 大矢野 is Wakamiya Swimming Monuments Amakusa u Campground Tourism Association Oyano r Area, Campground -

Kumamoto 5.7.17
KUMAMOTO, KYUSHU, JAPAN, EARTHQUAKES OF Mw 6.0 April 14, 2016 Mw 7.0 April 16, 2016 LIFELINE PERFORMANCE By ALEX K TANG and JOHN M EIDINGER The Council of Lifeline Earthquake Engineering TCLEE No. 2 and Yokohama National University May 7 2017 http://www.geEngineeringSystems.com Kumamoto Earthquakes April 2016 Rev. 0 April 16 2017 Table of Contents TABLE OF CONTENTS ............................................................................................................................... I ABSTRACT .................................................................................................................................................... 1 PREFACE ...................................................................................................................................................... 3 AUTHORS’ AFFILIATIONS ...................................................................................................................... 4 ACKNOWLEDGEMENTS .......................................................................................................................... 5 ENDORSEMENTS ........................................................................................................................................ 8 1.0 INTRODUCTION ................................................................................................................................... 9 1.1 LIMITATIONS .......................................................................................................................................