Mulualem Tamiru.Pdf
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-
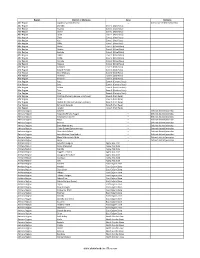
Districts of Ethiopia
Region District or Woredas Zone Remarks Afar Region Argobba Special Woreda -- Independent district/woredas Afar Region Afambo Zone 1 (Awsi Rasu) Afar Region Asayita Zone 1 (Awsi Rasu) Afar Region Chifra Zone 1 (Awsi Rasu) Afar Region Dubti Zone 1 (Awsi Rasu) Afar Region Elidar Zone 1 (Awsi Rasu) Afar Region Kori Zone 1 (Awsi Rasu) Afar Region Mille Zone 1 (Awsi Rasu) Afar Region Abala Zone 2 (Kilbet Rasu) Afar Region Afdera Zone 2 (Kilbet Rasu) Afar Region Berhale Zone 2 (Kilbet Rasu) Afar Region Dallol Zone 2 (Kilbet Rasu) Afar Region Erebti Zone 2 (Kilbet Rasu) Afar Region Koneba Zone 2 (Kilbet Rasu) Afar Region Megale Zone 2 (Kilbet Rasu) Afar Region Amibara Zone 3 (Gabi Rasu) Afar Region Awash Fentale Zone 3 (Gabi Rasu) Afar Region Bure Mudaytu Zone 3 (Gabi Rasu) Afar Region Dulecha Zone 3 (Gabi Rasu) Afar Region Gewane Zone 3 (Gabi Rasu) Afar Region Aura Zone 4 (Fantena Rasu) Afar Region Ewa Zone 4 (Fantena Rasu) Afar Region Gulina Zone 4 (Fantena Rasu) Afar Region Teru Zone 4 (Fantena Rasu) Afar Region Yalo Zone 4 (Fantena Rasu) Afar Region Dalifage (formerly known as Artuma) Zone 5 (Hari Rasu) Afar Region Dewe Zone 5 (Hari Rasu) Afar Region Hadele Ele (formerly known as Fursi) Zone 5 (Hari Rasu) Afar Region Simurobi Gele'alo Zone 5 (Hari Rasu) Afar Region Telalak Zone 5 (Hari Rasu) Amhara Region Achefer -- Defunct district/woredas Amhara Region Angolalla Terana Asagirt -- Defunct district/woredas Amhara Region Artuma Fursina Jile -- Defunct district/woredas Amhara Region Banja -- Defunct district/woredas Amhara Region Belessa -- -

John Percival
THE LINNEAN Wheat Taxonomy: the legacy of John Percival THE LINNEAN SOCIETY OF LONDON BURLINGTON HOUSE, PICCADILLY, LONDON WlJ OBF SPECIAL ISSUE No 3 2001 ACADEMIC PRESS LIMITED 32 Jam.estown Road London NWl 7BY Printed on acid free paper © 2001 The Linnean Society of London All rights reserved. No part of this book may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopy, recording, or any information storage or retrieval system without permission in writing from the publisher. The designations of geographic entities in this book, and the presentation of the material, do not imply the expression of any opinion whatsoever on the part of the publishers, the Linnean Society, the editors or any other participating organisations concerning the legal status of any country, territory, or area, or of its authorities, or concerning the delimitation of its frontiers or boundaries. The views expressed in this publication do not necessarily reflect those of The Society, the editors, or other participating organisations. Printed in Great Britain. Wheat Taxonomy: the legacy of John Percival Conference Participants (most of whom are identified by number on the key to the group photograph above). I. M. Ambrose,; 2. J. Bingham, UK; 3. R. Blatter, Switzerland; 4. A. Bomer, Germany; 5. A. Brandolini Italy; 6. R. Brigden, UK; 7. A. H. Bunting, UK; 8. P. Caligari, UK; 9. E.M.L.P. Clauss, USA; 10. P.O. Clauss, USA; 11 . K. Clavel, France; 12. P. Davis, UK; 13. J. Dvohik, USA; 14. !. Faberova, Czech Republic; 15 . A. A. Filatenko, Russia; 16. -

Improvement of Australian Wheat Grain Functionality for Breadmaking by Introgression of Novel High-Molecular Weight Glutenin Subunits Into Australian Cultivars
Improvement of Australian wheat grain functionality for breadmaking by introgression of novel high-molecular weight glutenin subunits into Australian cultivars NANDITA ROY A thesis submitted to Murdoch University in fulfilment of the requirements for the degree of Doctor of Philosophy Australia-China Joint Centre for Wheat Improvement School of Veterinary and Life Sciences Murdoch University Perth, Western Australia 2019 Declaration I am Nandita Roy, certify that: This work contains no material previously submitted for a degree or diploma in any University or other tertiary institution and, to the best of my knowledge and belief, no material which has been published or written by any other person except where due reference is made in the text. i Table of Contents Declaration i Table of Contents ii Acknowledgments x Abstract xii Abbreviations xiv Publications and conferences xvi List of Tables xvii List of Figures xviii Chapter 1 1 General Introduction 1 1.1 Aims of the project 1 1.2 Background 1 1.3 Thesis outline 3 1.4 References 4 Chapter 2 6 Literature review 6 2.1 Wheat 6 2.1.1 Evolution of bread wheat 6 2.1.2 Australian Wheat 7 2.1.3 Grain composition of wheat 7 ii 2.1.4 Storage protein and baking quality 8 2.2 Wheat grain protein composition and classification 10 2.2.1 Albumins 11 2.2.2 Globulins 11 2.2.3 Gluten 11 2.2.4 Gliadins 12 2.2.5 Glutenins 12 2.3 The genetics of gluten (gliadins and glutenins) 14 2.3.1 x and y-type HMW-GSs 15 2.3.2 Expression pattern of HMW glutenins genes in different types of wheat 16 2.4 The role of -

Enid Perez Lara
Mapping of genomic regions associated with agronomic traits and resistance to diseases in Canadian spring wheat by Enid Perez Lara A thesis submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy in Plant Science Department of Agricultural, Food and Nutritional Science University of Alberta © Enid Perez Lara, 2017 Abstract Wheat breeders, in addition to phenotypic selection, employ molecular markers in their programs for different purposes, including parental selection, quality control, analysis of advanced lines (cultivars), on genetic purity and identity, and for markers assisted selection. In the first study of this thesis we evaluated 158 recombinant inbred lines (RILs) population for flowering, maturity, plant height and grain yield under field conditions. With a subset of 1809 single nucleotide polymorphisms (SNPs) and 2 functional markers (Ppd-D1 and Rht-D1) we identified a total of 19 quantitative trait loci (QTLs) associated with flowering time under greenhouse (5) and field (6) conditions, maturity (5), grain yield (2) and plant height (1). These QTLs explained between 6.3 and 37.8% of the phenotypic variation. Only the QTLs on both 2D chromosome (adjacent to Ppd-D1) and 4D chromosome (adjacent Rht-D1) had major effects and, respectively reduced flowering and maturity time up to 5 days with a yield penalty of 436 kg ha-1 and reduced plant height by 13 cm, but increased maturity by 33 degree days. In the second study, we used genome-wide association analysis (GWAS) to identify markers associated with the wheat diseases leaf rust, stripe rust, tan spot, common bunt and three host selective toxins (HST) from Pyrenophora tritici-repentis (Ptr ToxA, B and C). -

Genetic and Genomic Analyses of Herbicide Tolerance in Bread Wheat (Triticum Aestivum L.)
Genetic and genomic analyses of herbicide tolerance in bread wheat (Triticum aestivum L.) Roopali Bhoite M.Tech (Biotechnology) B.Tech (Biotechnology) This thesis is presented for the degree of Doctor of Philosophy of The University of Western Australia UWA School of Agriculture and Environment Faculty of Science The University of Western Australia August 2019 THESIS DECLARATION I, Roopali Bhoite, certify that: This thesis has been substantially accomplished during enrolment in this degree. This thesis does not contain material which has been submitted for the award of any other degree or diploma in my name, in any university or other tertiary institution. In the future, no part of this thesis will be used in a submission in my name, for any other degree or diploma in any university or other tertiary institution without the prior approval of The University of Western Australia and where applicable, any partner institution responsible for the joint-award of this degree. This thesis does not contain any material previously published or written by another person, except where due reference has been made in the text and, where relevant, in the Authorship Declaration that follows. This thesis does not violate or infringe any copyright, trademark, patent, or other rights whatsoever of any person. This thesis contains published work and/or work prepared for publication, some of which has been co-authored. Signature: Date: 14/08/2019 ii ABSTRACT Wheat (Triticum aestivum L.) is one of the major global cereal grains in terms of production and area coverage (FAO 2018) and is Australia’s largest grain crop contributing up to 15% world trade. -

College of Social Science and Humanities Department of Geography and Environmental Studies
COLLEGE OF SOCIAL SCIENCE AND HUMANITIES DEPARTMENT OF GEOGRAPHY AND ENVIRONMENTAL STUDIES IMPACTS OF CLIMATE VARIABILITY ON SMALLHOLDER DAIRY FARMERS (THE CASE Of BASSONA WORANA WOREDA, NORTH SHEWA ZONE OF AMHARA NATIONAL REGIONAL STATE, ETHIOPIA) MSc. Thesis Tsegaye Hailiye July, 2020 Debre Berhan, Ethiopia i DEBRE BERHAN UNIVERSITY COLLEGE OF SOCIAL SCIENCE AND HUMANITIES DEPARTMENT OF GEOGRAPHY AND ENVIRONMENTAL STUDIES IMPACTS OF CLIMATE VARIABILITY ON SMALLHOLDER DAIRY FARMERS (THE CASE OF BASSONA WORANA WOREDA, NORTH SHEWA ZONE OF AMHARA NATIONAL REGIONAL STATE, ETHIOPIA) A Thesis Submitted to the Department of Geography and Environmental Studies, College Social Science and Humanities, Debre Berhan University In Partial Fulfillment of the Requirement for the Degree of Master of Science in Environment and Sustainable Development By Tsegaye Hailiye Adviser: Arragaw Alemayehu (PhD) July, 2020 Debre Berhan, Ethiopia ii DEBRE BERHAN UNIVERSITY COLLEGE SOCIAL SCIENCE AND HUMANITIES APPROVAL SHEET – I This is to certify that the thesis entitled: Impacts of Climate Variability on Smallholder Dairy Farmers in Bassona Worana Woreda (District), North Shewa Zone of Amhara National Regional State, Ethiopia submitted in partial fulfillment of the requirements for the degree of Masters of Science with specialization in Environment and Sustainable Development of the Graduate Program of the Geography and Environmental Studies College of Social Science and Humanities , Debre Berhan University and is a record of original research carried out by Tsegaye Hailiye PGR 024/11 under my supervision, and no part of the thesis has been submitted for any other degree or diploma. The assistance and help received during the course of this investigation have been duly acknowledged. -

A Dissertation Submitted To
The Political Economy of Environmental Governance in Ethiopia: Exploring Trends, Actors and Drivers Alemayehu Assefa Assele A Dissertation Submitted to The Center for Environment and Development, College of Development Studies Presented in partial fulfillment of the requirements for the Degree of Doctor of Philosophy in Development Studies (Environment and Development) Addis Ababa University Addis Ababa, Ethiopia May 2019 The Political Economy of Environmental Governance in Ethiopia: Exploring Trends, Actors and Drivers Alemayehu Assefa Assele A Dissertation Submitted to The Center for Environment and Development, College of Development Studies Presented in partial fulfillment of the requirements for the Degree of Doctor of Philosophy in Development Studies (Environment and Development) Addis Ababa University Addis Ababa, Ethiopia May 2019 ii Declaration I, the undersigned, declare that this is my original work, has never been presented in this or any other University, and that all the resources and materials used for the dissertation have been fully acknowledged. Name: Alemayehu Assefa Assele Signature: ____________ Date: _________________ Place: Addis Ababa Date of submission ___________________ This dissertation has been submitted for examination with my approval as University supervisor: Main advisor’s name: Yohannes Aberra (PhD) Signature: _________________________ Date: ________________________ Co-advisor’s name: Dawit Diriba (PhD) Signature: _____________________ Date: ________________________ iii Dissertation Approval Addis Ababa University School of Graduate Studies This is to certify that the dissertation prepared by Alemayehu Assefa Assele entitled “The Political Economy of Environmental Governance in Ethiopia: Exploring Trends, Actors and Drivers” and submitted in fulfillment of the requirement for the Degree of Doctor of Philosophy (Environment and Development Studies) complies with the regulations of the University and meets the accepted standards with respect to originality and quality. -

Plant Genetics, Genomics, Bioinformatics and Biotechnology" (Plantgen2017)
PROCEEDINGS of the 4th International conference "Plant Genetics, Genomics, Bioinformatics and Biotechnology" (PlantGen2017) Best Western Plus Atakent Park Hotel May 29 – June 02, 2017, Almaty, Kazakhstan УДК 581 (063) ББК 28.5 Р71 "Plant Genetics, Genomics, Bioinformatics and Biotechnology": Материалы Международной конференции 4th International conference PlantGen2017 / под общей редакцией Е.К. Туруспекова, С.И. Абугалиевой. – Алматы: ИББР, 2017 – 216 с. ISBN 978-601-80631-2-1 В сборнике представлены материалы 4 Международной конференции по генетике, геномике, биоинформатике и биотехнологии растений (PlantGen2017), проведенной в г. Алматы 29 мая -2 июня 2017 г. В публикациях изложены результаты оригинальных исследований в области изучения, сохранения и использования генетических ресурсов, генетики и селекции, биоинформатики и биотехнологии растений. Сборник рассчитан на биологов, генетиков, биотехнологов, селекционеров, специалистов, занимающихся генетическими ресурсами растений, и студентов биологического и сельско-хозяйственного профиля. Тезисы докладов представлены в авторской редакции. Рекомендовано к изданию Ученым советом РГП «Института биологии и биотехнологии растений» Комитета науки Министерства образования и науки Республики Казахстан (Протокол № 2 от 04.05.2017 г.). УДК 581 (063) ББК 285 ISBN 978-601-80631-2-1 © ИББР, 2017 2 Proceedings of the 4th International Conference "Plant Genetics, Genomics, Bioinformatics and Biotechnology" (PlantGen2017) May 29 – June 02 2017 – Almaty, Kazakhstan Editors Yerlan Turuspekov, Saule Abugalieva Publisher Institute of Plant Biology and Biotechnology Plant Molecular Genetics Lab ISBN: 978-601-80631-2-1 Responsibility for the text content of each abstract is with the respective authors. Date: May 29 – June 02 2017 Venue: Best Western Plus Atakent Park Hotel, 42 Timiryazev str., 050040 Almaty, Kazakhstan Conference webpage: http://primerdigital.com/PlantGen2017/en/ Hosted by: Institute of Plant Biology and Biotechnology (IPBB), Almaty, Kazakhstan Correct citation: Turuspekov Ye., Abugalieva S. -

The Contribution of Ethiopian Orthodox Tewahido Church in Forest Management and Its Best Practices to Be Scaled up in North Shewa Zone of Amhara Region, Ethiopia
Agriculture, Forestry and Fisheries 2015; 4(3): 123-137 Published online June 4, 2015 (http://www.sciencepublishinggroup.com/j/aff) doi: 10.11648/j.aff.20150403.18 ISSN:2328-563X (Print); ISSN:2328-5648 (Online) The Contribution of Ethiopian Orthodox Tewahido Church in Forest Management and Its Best Practices to be Scaled up in North Shewa Zone of Amhara Region, Ethiopia Abiyou Tilahun 1, *, Hailu Terefe 1, Teshome Soromessa 2 1Department of Biology, College of Natural Science, Debre Berhan University, Debre Berhan, Ethiopia 2Department of Environmental Science, Science faculty, Addis Ababa University, Addis Ababa, Ethiopia Email address: [email protected] (A. Tilahun) To cite this article: Abiyou Tilahun, Hailu Terefe, Teshome Soromessa. The Contribution of Ethiopian Orthodox Tewahido Church in Forest Management and Its Best Practices to be Scaled up in North Shewa Zone of Amhara Region, Ethiopia. Agriculture, Forestry and Fisheries . Vol. 4, No. 3, 2015, pp. 123-137. doi: 10.11648/j.aff.20150403.18 Abstract: This research was conducted in selected moasteries of Ethiopian Orthodox Church (EOTC) in North Shewa zone. The main objective of the study is to identify main constraints which hinder the society to learn from religious instituions and apply the best practices and habit of experiences on forest conservation. Accordingly six monasteries were selected at different altitudes and sites purposively. Once the study forest areas were identified, 10 X 10 m quadrats were laid systematically in the forests for vegetation sampling. For the socioeconomic survey, 112 individuals were selected. Moreover, focus group discussion and key informant interviews were employed. Church forests enveloped in this study have an area ranging from 1.6 ha to 100 ha. -

Addis Ababa University School of Graduate Studies Department of Sociology
ADDIS ABABA UNIVERSITY SCHOOL OF GRADUATE STUDIES DEPARTMENT OF SOCIOLOGY THE ROLE OF AMHARA CREDIT AND SAVING INSTITUTION (ACSI) IN THE SOCIO-ECONOMIC EMPOWERMENT OF WOMEN IN BASONA WERANA WOREDA BY DAGNACHEW TEREFE JUNE 2019 ADDIS ABABA, ETHIOPIA Addis Ababa University School of Graduate Studies Department of Sociology The Role of Amhara Credit and Saving Institution (ACSI) in the Socio- Economic Empowerment of Women in Basona Werana Woreda A Thesis Submitted to the School of Graduate Studies of Addis Ababa University in Partial Fulfillment of the Requirements for the Degree of Master of Arts in Sociology By Dagnachew Terefe Advisor Woldeab Teshome (PhD) June 2019 Addis Ababa, Ethiopia Declaration I hereby declare that the thesis entitled: ―The Role of Amhara Credit and Saving Institution (ACSI) in the Socio-Economic Empowerment of Women in Basona Werana Woreda‖, submitted to award of the Degree of Master of Arts in Sociology at Addis Ababa University, is my original research work that has not been presented anywhere for any degree. To the best of the researcher‘s knowledge, all sources of material used for the thesis have been fully acknowledged. Name: Dagnachew Terefe Signature: ______________ Date: June 2019 i ADDIS ABABA UNIVERSITY SCHOOL OF GRADUATE STUDIES This is to certify that the thesis prepared by Dagnachew Terefe, entitled: The Role of Amhara Credit and Saving Institution (ACSI) in the Socio-Economic Empowerment of Women in Basona Werana Woreda and submitted in partial fulfillment of the requirements for the Degree of Master of Arts in Sociology complies with the regulations of the University and meets the accepted standards with respect to originality and quality. -

Phenolic Acids and Antioxidative Capacity on Ancient Wheat Namely Einkorn (T
Justus Liebig University Gießen Faculty of Agricultural Sciences, Nutritional Sciences and Environmental Management Institute of Agronomy and Plant Breeding I Head: Prof. Dr. Bernd Honermeier Phenolic acids and antioxidative capacity on ancient wheat namely einkorn (T. monococcum ssp.), emmer (T. turgidum ssp.) and spelt wheat (T. aestivum ssp. spelta) and on germinated bread wheat (T. aestivum ssp. aestivum) A dissertation submitted for the degree of Doctor of home economics and nutritional science (Doctor oeconomiae trophologiaeque- Dr. oec. troph.) in the Faculty of Agricultural Sciences, Nutritional Sciences, and Environmental Management at Justus Liebig University Gießen Submitted by: Nadine Engert, M. Sc. From Glauchau Gießen, June 24, 2011 Phenolic compounds: Are they healthy? Are they present in wheat? Have they always been present in wheat? Fig. 0-1:Health benefits by whole grain consumption (VITAGLIONE et al., 2008) II Table of Contents Phenolic compounds: II Index of Tables V Index of Figures VII Index of Equations IX Abbreviations X 1 Introduction 1 2 Objectives 6 3 Literature review 8 3.1 Characterization of the genus Triticum 8 3.1.1 Domestication 8 3.1.2 Taxonomy of wheat 10 3.2 Secondary plant metabolites 12 3.2.1 Phenolic acids 14 3.2.2 Oxidative stress 18 3.2.3 Health effecting potentials and nutrition of phenolic compounds 23 3.2.4 Bioavailability 32 3.3 Phenolic acids in wheat 35 3.4 Antioxidative capacity tests 36 4 Material and methods 53 4.1 Characterization of the experiments 53 4.2 Grain quality parameters 55 -

Plant Diversity of Mediterranean-Type Ecosystems with an Emphasis on Cultivated Species
PLANT DIVERSITY OF MEDITERRANEAN-TYPE ECOSYSTEMS WITH AN EMPHASIS ON CULTIVATED SPECIES by Mina Habib Botros A professional paper submitted in partial fulfillment of the requirements for the degree of Master of Science in Plant Sciences MONTANA STATE UNIVERSITY Bozeman, Montana November 2019 ©COPYRIGHT by Mina Habib Botros 2019 All Rights Reserved ii DEDICATION To my parents and my brother “Remon” who have supported me throughout my graduate studies. May God bless them. iii ACKNOWLEDGEMENTS I would like to express my gratitude and respect to my supervisor Dr. Matthew Lavin for his scientific guidance, supervision, support, and his constructive attitude throughout the preparation of this degree. I have been extremely lucky to have the opportunity to work with an experienced scientist. Many thanks go in particular to the committee members: Dr. Luther Talbert and Dr. Norman Weeden for their supervision, and crucial contribution to this project; I appreciate their time and support. Also, I would like to thank Dr. George Haynes and Dr. Carl Igo for the teaching experience I gained through working with them. Thanks go to the department of Plant Sciences & Plant Pathology, headed by Dr. Michael Giroux, for providing the financial support and facilities to accomplish this degree. My thanks go to Montana State University writing center for the English editing of this study and for being a wonderful help. I would like to express my deep sincere love and appreciation to my lab and department colleagues for their support, cooperation, and the pleasant time we spent working together. Warm thanks go to all my friends for creating such a pleasant atmosphere which made my stay in Bozeman easier, enjoyable, and an everlasting nice memory.