Zootaxa, Elachista (Lepidoptera: Gelechioidea: Elachistidae)
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Self-Repair and Self-Cleaning of the Lepidopteran Proboscis
Clemson University TigerPrints All Dissertations Dissertations 8-2019 Self-Repair and Self-Cleaning of the Lepidopteran Proboscis Suellen Floyd Pometto Clemson University, [email protected] Follow this and additional works at: https://tigerprints.clemson.edu/all_dissertations Recommended Citation Pometto, Suellen Floyd, "Self-Repair and Self-Cleaning of the Lepidopteran Proboscis" (2019). All Dissertations. 2452. https://tigerprints.clemson.edu/all_dissertations/2452 This Dissertation is brought to you for free and open access by the Dissertations at TigerPrints. It has been accepted for inclusion in All Dissertations by an authorized administrator of TigerPrints. For more information, please contact [email protected]. SELF-REPAIR AND SELF-CLEANING OF THE LEPIDOPTERAN PROBOSCIS A Dissertation Presented to the Graduate School of Clemson University In Partial Fulfillment of the Requirements for the Degree Doctor of Philosophy ENTOMOLOGY by Suellen Floyd Pometto August 2019 Accepted by: Dr. Peter H. Adler, Major Advisor and Committee Co-Chair Dr. Eric Benson, Committee Co-Chair Dr. Richard Blob Dr. Patrick Gerard i ABSTRACT The proboscis of butterflies and moths is a key innovation contributing to the high diversity of the order Lepidoptera. In addition to taking nectar from angiosperm sources, many species take up fluids from overripe or sound fruit, plant sap, animal dung, and moist soil. The proboscis is assembled after eclosion of the adult from the pupa by linking together two elongate galeae to form one tube with a single food canal. How do lepidopterans maintain the integrity and function of the proboscis while foraging from various substrates? The research questions included whether lepidopteran species are capable of total self- repair, how widespread the capability of self-repair is within the order, and whether the repaired proboscis is functional. -

SYSTEMATICS of the MEGADIVERSE SUPERFAMILY GELECHIOIDEA (INSECTA: LEPIDOPTEA) DISSERTATION Presented in Partial Fulfillment of T
SYSTEMATICS OF THE MEGADIVERSE SUPERFAMILY GELECHIOIDEA (INSECTA: LEPIDOPTEA) DISSERTATION Presented in Partial Fulfillment of the Requirements for The Degree of Doctor of Philosophy in the Graduate School of The Ohio State University By Sibyl Rae Bucheli, M.S. ***** The Ohio State University 2005 Dissertation Committee: Approved by Dr. John W. Wenzel, Advisor Dr. Daniel Herms Dr. Hans Klompen _________________________________ Dr. Steven C. Passoa Advisor Graduate Program in Entomology ABSTRACT The phylogenetics, systematics, taxonomy, and biology of Gelechioidea (Insecta: Lepidoptera) are investigated. This superfamily is probably the second largest in all of Lepidoptera, and it remains one of the least well known. Taxonomy of Gelechioidea has been unstable historically, and definitions vary at the family and subfamily levels. In Chapters Two and Three, I review the taxonomy of Gelechioidea and characters that have been important, with attention to what characters or terms were used by different authors. I revise the coding of characters that are already in the literature, and provide new data as well. Chapter Four provides the first phylogenetic analysis of Gelechioidea to include molecular data. I combine novel DNA sequence data from Cytochrome oxidase I and II with morphological matrices for exemplar species. The results challenge current concepts of Gelechioidea, suggesting that traditional morphological characters that have united taxa may not be homologous structures and are in need of further investigation. Resolution of this problem will require more detailed analysis and more thorough characterization of certain lineages. To begin this task, I conduct in Chapter Five an in- depth study of morphological evolution, host-plant selection, and geographical distribution of a medium-sized genus Depressaria Haworth (Depressariinae), larvae of ii which generally feed on plants in the families Asteraceae and Apiaceae. -

Download PDF ( Final Version , 576Kb )
sys: 7<3èV?ïl Bd £ß) f ENTOMOLOGISCHE MAANDBLAD UITGEGEVEN DOOR DE NEDERLANDSE ENTOMOLOGISCHE VERENIGING Officiële afkorting (World List): Ent. Ber., Amst. Deel 35 1 augustus 1975 No. 8 Adres van de Redactie: B. J. LEMPKE, Plantage Middenlaan 64, Amsterdam 1004 — Nederland INHOUD: B. J. LEMPKE, Notities over enkele soorten van Elachista (Lep., Elachisti- dae) (p. 105). — A. VAN FRANKENHUYZEN, Phyllonorycter corylifoliella (Hübner, 1793) (Lep., Gracillariidae) (p. 108). — FER WILLEMSE, Two new subspecies of Platycleis (Modestana) ebneri (Ramme, 1926) from Greece (Orthoptera, Ensifera, Dec- ticinae) (p. 118). — Literatuur (p. 124: A. DIAKONOFF). — Korte mededeling (p. 124: Personalia). Notities over enkele soorten van Elachista (Lep., Elachistidae) door B. J. LEMPKE In 1867 ving Van Medenbach de Rooij bij Arnhem een Elachista met lichtgele kop, terwijl de voorvleugels eenkleurig zwart zijn met geelachtige middenband. Snellen stuurde het nogal afgevlogen exemplaar naar Zeller, die het determineerde als Elachista cinctella. Onder deze naam met Zeiler als auteur publiceerden Snellen en De Graaf de vangst (1869: 211). Na het eerste exemplaar ving Van Medenbach nog enkele andere, voor zover bekend het laatst in 1872. Vier daarvan bevinden zich in de collectie van het Amsterdams museum, een vijfde in de collectie-Bentinck te Leiden. De data liggen tussen 1 en 23 juli. In zijn standaardwerk over de Nederlandse Microlepidoptera (1882: 884) noemde Snellen de soort megerlella Tengström, 1848, met adscitella Stainton en abruptella Stainton als synoniemen. Rebel (1901: 203, nr. 3978) tenslotte noemde de soort megerlella Stainton met cinctella Zeiler, adscitella Stainton en obliquella Stainton als synoniemen en onder deze naam komt de soort ook voor in de Naamlijst van Lycklama (1925, 1927). -

Hym.: Eulophidae) New Larval Ectoparasitoids of Tuta Absoluta (Meyreck) (Lep.: Gelechidae)
J. Crop Prot. 2016, 5 (3): 413-418______________________________________________________ Research Article Two species of the genus Elachertus Spinola (Hym.: Eulophidae) new larval ectoparasitoids of Tuta absoluta (Meyreck) (Lep.: Gelechidae) Fatemeh Yarahmadi1*, Zohreh Salehi1 and Hossein Lotfalizadeh2 1. Ramin Agriculture and Natural Resources University, Mollasani, Ahvaz, Iran. 2. East-Azarbaijan Research Center for Agriculture and Natural Resources, Tabriz, Iran. Abstract: This is the first report of two ectoparasitoid wasps, Elachertus inunctus (Nees, 1834) in Iran and Elachertus pulcher (Erdös, 1961) (Hym.: Eulophidae) in the world, that parasitize larvae of the tomato leaf miner, Tuta absoluta (Meyrick, 1917) (Lep.: Gelechiidae). The specimens were collected from tomato fields and greenhouses in Ahwaz, Khouzestan province (south west of Iran). Both species are new records for fauna of Iran. The knowledge about these parasitoids is still scanty. The potential of these parasitoids for biological control of T. absoluta in tomato fields and greenhouses should be investigated. Keywords: tomato leaf miner, parasitoids, identification, biological control Introduction12 holometabolous insects, the overall range of hosts and biologies in eulophid wasps is remarkably The Eulophidae is one of the largest families of diverse (Gauthier et al., 2000). Chalcidoidea. The chalcid parasitoid wasps attack Species of the genus Elachertus Spinola, 1811 insects from many orders and also mites. Many (Hym.: Eulophidae) are primary parasitoids of a eulophid wasps parasitize several pests on variety of lepidopteran larvae. Some species are different crops. They can regulate their host's polyphagous that parasite hosts belonging to populations in natural conditions (Yefremova and different insect families. The larvae of these Myartseva, 2004). Eulophidae are composed of wasps are often gregarious and their pupae can be four subfamilies, Entedoninae (Förster, 1856), observed on the surface of plant leaves or the Euderinae (Lacordaire, 1866), Eulophinae body of their host. -

Lepidoptera of North America 5
Lepidoptera of North America 5. Contributions to the Knowledge of Southern West Virginia Lepidoptera Contributions of the C.P. Gillette Museum of Arthropod Diversity Colorado State University Lepidoptera of North America 5. Contributions to the Knowledge of Southern West Virginia Lepidoptera by Valerio Albu, 1411 E. Sweetbriar Drive Fresno, CA 93720 and Eric Metzler, 1241 Kildale Square North Columbus, OH 43229 April 30, 2004 Contributions of the C.P. Gillette Museum of Arthropod Diversity Colorado State University Cover illustration: Blueberry Sphinx (Paonias astylus (Drury)], an eastern endemic. Photo by Valeriu Albu. ISBN 1084-8819 This publication and others in the series may be ordered from the C.P. Gillette Museum of Arthropod Diversity, Department of Bioagricultural Sciences and Pest Management Colorado State University, Fort Collins, CO 80523 Abstract A list of 1531 species ofLepidoptera is presented, collected over 15 years (1988 to 2002), in eleven southern West Virginia counties. A variety of collecting methods was used, including netting, light attracting, light trapping and pheromone trapping. The specimens were identified by the currently available pictorial sources and determination keys. Many were also sent to specialists for confirmation or identification. The majority of the data was from Kanawha County, reflecting the area of more intensive sampling effort by the senior author. This imbalance of data between Kanawha County and other counties should even out with further sampling of the area. Key Words: Appalachian Mountains, -

The Biology of Casmara Subagronoma (Lepidoptera
insects Article The Biology of Casmara subagronoma (Lepidoptera: Oecophoridae), a Stem-Boring Moth of Rhodomyrtus tomentosa (Myrtaceae): Descriptions of the Previously Unknown Adult Female and Immature Stages, and Its Potential as a Biological Control Candidate Susan A. Wineriter-Wright 1, Melissa C. Smith 1,* , Mark A. Metz 2 , Jeffrey R. Makinson 3 , Bradley T. Brown 3, Matthew F. Purcell 3, Kane L. Barr 4 and Paul D. Pratt 5 1 USDA-ARS Invasive Plant Research Laboratory, Fort Lauderdale, FL 33314, USA; [email protected] 2 USDA-ARS Systematic Entomology Lab, Beltsville, MD 20013-7012, USA; [email protected] 3 USDA-ARS Australian Biological Control Laboratory, CSIRO Health and Biosecurity, Dutton Park QLD 4102, Australia; jeff[email protected] (J.R.M.); [email protected] (B.T.B.); [email protected] (M.F.P.) 4 USDA-ARS Center for Medical, Agricultural and Veterinary Entomology, Gainesville, FL 32608, USA; [email protected] 5 USDA-ARS, Western Regional Research Center, Invasive Species and Pollinator Health Research Unit, 800 Buchanan Street, Albany, CA 94710, USA; [email protected] * Correspondence: [email protected]; Tel.: +1-954-475-6549 Received: 27 August 2020; Accepted: 16 September 2020; Published: 23 September 2020 Simple Summary: Rhodomyrtus tomentosa is a perennial woody shrub throughout Southeast Asia. Due to its prolific flower and fruit production, it was introduced into subtropical areas such as Florida and Hawai’i, where it is now naturalized and invasive. In an effort to find sustainable means to control R. tomentosa, a large-scale survey was mounted for biological control organisms. -

Big Creek Lepidoptera Checklist
Big Creek Lepidoptera Checklist Prepared by J.A. Powell, Essig Museum of Entomology, UC Berkeley. For a description of the Big Creek Lepidoptera Survey, see Powell, J.A. Big Creek Reserve Lepidoptera Survey: Recovery of Populations after the 1985 Rat Creek Fire. In Views of a Coastal Wilderness: 20 Years of Research at Big Creek Reserve. (copies available at the reserve). family genus species subspecies author Acrolepiidae Acrolepiopsis californica Gaedicke Adelidae Adela flammeusella Chambers Adelidae Adela punctiferella Walsingham Adelidae Adela septentrionella Walsingham Adelidae Adela trigrapha Zeller Alucitidae Alucita hexadactyla Linnaeus Arctiidae Apantesis ornata (Packard) Arctiidae Apantesis proxima (Guerin-Meneville) Arctiidae Arachnis picta Packard Arctiidae Cisthene deserta (Felder) Arctiidae Cisthene faustinula (Boisduval) Arctiidae Cisthene liberomacula (Dyar) Arctiidae Gnophaela latipennis (Boisduval) Arctiidae Hemihyalea edwardsii (Packard) Arctiidae Lophocampa maculata Harris Arctiidae Lycomorpha grotei (Packard) Arctiidae Spilosoma vagans (Boisduval) Arctiidae Spilosoma vestalis Packard Argyresthiidae Argyresthia cupressella Walsingham Argyresthiidae Argyresthia franciscella Busck Argyresthiidae Argyresthia sp. (gray) Blastobasidae ?genus Blastobasidae Blastobasis ?glandulella (Riley) Blastobasidae Holcocera (sp.1) Blastobasidae Holcocera (sp.2) Blastobasidae Holcocera (sp.3) Blastobasidae Holcocera (sp.4) Blastobasidae Holcocera (sp.5) Blastobasidae Holcocera (sp.6) Blastobasidae Holcocera gigantella (Chambers) Blastobasidae -

Elachista Saccharella (Lepidoptera: Elachistidae), a Leafminer Infesting Sugarcane in Louisiana
792 Florida Entomologist 90(4) December 2007 ELACHISTA SACCHARELLA (LEPIDOPTERA: ELACHISTIDAE), A LEAFMINER INFESTING SUGARCANE IN LOUISIANA W. H. WHITE1, T. E. REAGAN2, C. CARLTON2, W. AKBAR2 AND J. M. BEUZELIN2 1USDA, ARS, Sugarcane Research Laboratory, Houma, LA 2Department of Entomology, Louisiana State University Agricultural Center, Baton Rouge, LA A leafminer was discovered infesting sugar- Elachista saccharella was first reported in sug- cane (interspecific hybrids of Saccharum spp.) in arcane in the U.S. in Florida in 1982 (Hall 1984). Terrebonne Parish (near Schriever, LA, 29.38°N, It was previously reported in Cuba by Scara- 90.50°W) during the summer of 2006. Larvae muzza & Barry (1959). An unidentified elachistid were collected from the field, returned to the lab- also was found in sugarcane in Papua New oratory, and placed on sugarcane borer, Diatraea Guinea by Bourke (1968). Elachista saccharella saccharalis (F.), meridic diet (Southland Products, also has been reported in reed grass, Phragmites Lake Village, AR). Emerged adults were identified australis (Cav) Trin ex Steud, a plant invader of as Elachista saccharella (Busck) (Lepidoptera: salt marshes along the east coast of North Amer- Elachistidae) (Fig. 1a). Elachista saccharella is a ica (Gratton & Denno 2005). blotch leafminer. Each leaf mine begins as a longi- We initiated a survey of sugarcane growing ar- tudinal mine on the underside of a sugarcane leaf eas of Louisiana on 16 Aug 2006. Twenty parishes that sometimes extends 12 cm or more in length were surveyed by sampling 4 sites per parish for and terminates in a leaf blotch (Fig. 1b). These the presence of the leafminer. -
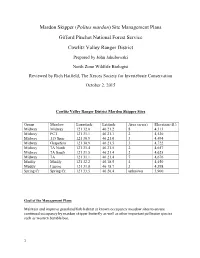
Mardon Skipper Site Management Plans
Mardon Skipper (Polites mardon) Site Management Plans Gifford Pinchot National Forest Service Cowlitz Valley Ranger District Prepared by John Jakubowski North Zone Wildlife Biologist Reviewed by Rich Hatfield, The Xerces Society for Invertebrate Conservation October 2, 2015 Cowlitz Valley Ranger District Mardon Skipper Sites Group Meadow Longitude Latitude Area (acres) Elevation (ft.) Midway Midway 121 32.0 46 21.2 8 4,313 Midway PCT 121 31.1 46 21.1 2 4,530 Midway 115 Spur 121 30.9 46 21.0 3 4,494 Midway Grapefern 121 30.9 46 21.5 3 4,722 Midway 7A North 121 31.4 46 21.5 2 4,657 Midway 7A South 121 31.5 46 21.4 2 4,625 Midway 7A 121 31.1 46 21.4 7 4,676 Muddy Muddy 121 32.2 46.18.5 4 4,450 Muddy Lupine 121 31.8 46 18.7 3 4,398 Spring Cr Spring Cr. 121 33.5 46 20.4 unknown 3,900 Goal of the Management Plans Maintain and improve grassland/forb habitat at known occupancy meadow sites to ensure continued occupancy by mardon skipper butterfly as well as other important pollinator species such as western bumble bee. 1 Introduction On the Gifford Pinchot National Forest (GPNF), mardon skippers were first detected on the Mt. Adams Ranger District (MTA) in 2000 and on Cowlitz Valley Ranger District (CVRD) in 2002. Mardon skippers are known to inhabit ten, upland dry grassy meadows on the CVRD. Portions of the meadows are mesic and are unsuitable mardon skipper habitat. -

Definition of the Elachista Puplesisi Sruoga Complex (Lepidoptera, Gelechioidea, Elachistidae), with Description of a New Species
Zootaxa 3821 (5): 583–589 ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ Article ZOOTAXA Copyright © 2014 Magnolia Press ISSN 1175-5334 (online edition) http://dx.doi.org/10.11646/zootaxa.3821.5.6 http://zoobank.org/urn:lsid:zoobank.org:pub:94D1D19B-9372-4DFC-985A-A4F450023F56 Definition of the Elachista puplesisi Sruoga complex (Lepidoptera, Gelechioidea, Elachistidae), with description of a new species LAURI KAILA1,3 & VIRGINIJUS SRUOGA2 1Finnish Museum of Natural History, Zoology Unit, FI-00014 University of Helsinki, Finland. E-mail: [email protected] 2Division of Biosystematics Research, Department of Biology, Lithuanian University of Educational Sciences, LT-08106 Vilnius, Lit- huania. E-mail: [email protected] 3Corresponding author Abstract The Elachista puplesisi group is defined. Its characterization is based on two species, E. puplesisi Sruoga, 2000, known from the holotype collected in Turkmenistan and E. helia sp. nov., from Rhodos, Greece. The group is assigned to the subgenus Atachia of Elachista, but its affinities within Atachia remain unknown. Key words: Elachistinae, Elachista, Atachia Introduction Elachista puplesisi Sruoga, 2000 was described on the basis of a single male specimen collected from the Karakum desert in Turkmenistan. The species appeared bizarre in its morphology, and its systematic position was left unresolved in the original description, with the remark that it likely represents a separate species group within Elachista, or even a new genus (Sruoga 2000). The species indeed possesses features rare among Elachista, as well as an unusual combination of characters, which did not make it straightforward to place this species in the phylogenetic concept of Elachistinae of Kaila (1999). -

Two New Records for the Appalachian Grizzled Skipper (Pyrgus Wyandot)
Banisteria, Number 24, 2004 © 2004 by the Virginia Natural History Society Status of the Appalachian Grizzled Skipper (Pyrgus centaureae wyandot) in Virginia Anne C. Chazal, Steven M. Roble, Christopher S. Hobson, and Katharine L. Derge1 Virginia Department of Conservation and Recreation Division of Natural Heritage 217 Governor Street Richmond, Virginia 23219 ABSTRACT The Appalachian grizzled skipper (Pyrgus centaureae wyandot) was documented historically (primarily from shale barren habitats) in 11 counties in Virginia. Between 1992 and 2002, staff of the Virginia Department of Conservation and Recreation, Division of Natural Heritage, conducted 175 surveys for P. c. wyandot at 75 sites in 12 counties. The species was observed at only six sites during these surveys, representing two new county records. All observations since 1992 combined account for <80 individuals. Due to forest succession and threats from gypsy moth control measures, all recent sites for P. c. wyandot in Virginia may be degrading in overall habitat quality. Key words: Lepidoptera, Pyrgus centaureae wyandot, conservation, shale barrens, Virginia. INTRODUCTION wyandot) in Virginia. Parshall (2002) provides a comprehensive review of the nomenclature and The Appalachian grizzled skipper (Pyrgus taxonomy of P. c. wyandot. Most authors classify this centaureae wyandot) has a rather fragmented range, skipper as a subspecies of the Holarctic Pyrgus occurring in northern Michigan as well as portions of centaureae (e.g., Opler & Krizek, 1984; Iftner et al., Ohio, Pennsylvania, Maryland, West Virginia, and 1992; Shuey, 1994; Allen, 1997; Opler, 1998; Virginia; isolated historical records are known from Glassberg, 1999; Parshall, 2002), although some Kentucky, New York, New Jersey, North Carolina, and lepidopterists treat it as a full species (Shapiro, 1974; the District of Columbia (Opler, 1998; NatureServe, Schweitzer, 1989; Gochfeld & Burger, 1997). -

Additions, Deletions and Corrections to An
Bulletin of the Irish Biogeographical Society No. 36 (2012) ADDITIONS, DELETIONS AND CORRECTIONS TO AN ANNOTATED CHECKLIST OF THE IRISH BUTTERFLIES AND MOTHS (LEPIDOPTERA) WITH A CONCISE CHECKLIST OF IRISH SPECIES AND ELACHISTA BIATOMELLA (STAINTON, 1848) NEW TO IRELAND K. G. M. Bond1 and J. P. O’Connor2 1Department of Zoology and Animal Ecology, School of BEES, University College Cork, Distillery Fields, North Mall, Cork, Ireland. e-mail: <[email protected]> 2Emeritus Entomologist, National Museum of Ireland, Kildare Street, Dublin 2, Ireland. Abstract Additions, deletions and corrections are made to the Irish checklist of butterflies and moths (Lepidoptera). Elachista biatomella (Stainton, 1848) is added to the Irish list. The total number of confirmed Irish species of Lepidoptera now stands at 1480. Key words: Lepidoptera, additions, deletions, corrections, Irish list, Elachista biatomella Introduction Bond, Nash and O’Connor (2006) provided a checklist of the Irish Lepidoptera. Since its publication, many new discoveries have been made and are reported here. In addition, several deletions have been made. A concise and updated checklist is provided. The following abbreviations are used in the text: BM(NH) – The Natural History Museum, London; NMINH – National Museum of Ireland, Natural History, Dublin. The total number of confirmed Irish species now stands at 1480, an addition of 68 since Bond et al. (2006). Taxonomic arrangement As a result of recent systematic research, it has been necessary to replace the arrangement familiar to British and Irish Lepidopterists by the Fauna Europaea [FE] system used by Karsholt 60 Bulletin of the Irish Biogeographical Society No. 36 (2012) and Razowski, which is widely used in continental Europe.