Habitat Selection and Partitioning Among Darters in Two Tributaries of the Clinch River, and Stream Restoration Effects on Substrate Profile
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Species Status Assessment Report for the Barrens Darter (Etheostoma Forbesi)
Species Status Assessment Report for the Barrens Darter (Etheostoma forbesi) Version 2.0 Acknowledgements: This Species Status Assessment would not have been possible without the research and assistance of Dr. Richard Harrington, Yale University Department of Ecology and Evolutionary Biology, Dr. Hayden Mattingly and his students, Tennessee Tech University School of Environmental Studies, Dr. John Johansen, Austin Peay State University Department of Biology, and Mark Thurman, Tennessee Wildlife Resources Agency. 1 TABLE OF CONTENTS Chapter 1: Introduction ............................................................................................................... 3 Chapter 2: Biology and Life History ........................................................................................... 4 Taxonomy ................................................................................................................................ 4 Genetic Diversity ..................................................................................................................... 5 Morphological Description ...................................................................................................... 5 Habitat ..................................................................................................................................... 6 Lifecycle .................................................................................................................................. 7 Population Needs .................................................................................................................... -

Percina Copelandi) in CANADA
Canadian Science Advisory Secretariat Central and Arctic, and Quebec Regions Science Advisory Report 2010/058 RECOVERY POTENTIAL ASSESSMENT OF CHANNEL DARTER (Percina copelandi) IN CANADA Channel Darter (Percina copelandi) Figure 1. Distribution of Channel Darter in Canada. © Ellen Edmondson Context : The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) assessed the status of Channel Darter (Percina copelandi) in April 1993. The assessment resulted in the designation of Channel Darter as Threatened. In May 2002, the status was re-examined and confirmed by COSEWIC. This designation was assigned because the species exists in low numbers where found, and its habitat is negatively impacted by siltation and fluctuations in water levels. Subsequent to the COSEWIC designation, Channel Darter was included on Schedule 1 of the Species at Risk Act (SARA) when the Act was proclaimed in June 2003. A species Recovery Potential Assessment (RPA) process has been developed by Fisheries and Oceans Canada (DFO) Science to provide the information and scientific advice required to meet the various requirements of the SARA, such as the authorization to carry out activities that would otherwise violate the SARA as well as the development of recovery strategies. The scientific information also serves as advice to the Minister of DFO regarding the listing of the species under SARA and is used when analyzing the socio-economic impacts of adding the species to the list as well as during subsequent consultations, where applicable. This assessment considers the scientific data available with which to assess the recovery potential of Channel Darter in Canada. SUMMARY In Ontario, the current and historic Channel Darter distribution is limited to four distinct areas of the Great Lakes basin: Lake St. -

Fish Inventory at Stones River National Battlefield
Fish Inventory at Stones River National Battlefield Submitted to: Department of the Interior National Park Service Cumberland Piedmont Network By Dennis Mullen Professor of Biology Department of Biology Middle Tennessee State University Murfreesboro, TN 37132 September 2006 Striped Shiner (Luxilus chrysocephalus) – nuptial male From Lytle Creek at Fortress Rosecrans Photograph by D. Mullen Table of Contents List of Tables……………………………………………………………………….iii List of Figures………………………………………………………………………iv List of Appendices…………………………………………………………………..v Executive Summary…………………………………………………………………1 Introduction…………………………………………………………………...……..2 Methods……………………………………………………………………………...3 Results……………………………………………………………………………….7 Discussion………………………………………………………………………….10 Conclusions………………………………………………………………………...14 Literature Cited…………………………………………………………………….15 ii List of Tables Table1: Location and physical characteristics (during September 2006, and only for the riverine sites) of sample sites for the STRI fish inventory………………………………17 Table 2: Biotic Integrity classes used in assessing fish communities along with general descriptions of their attributes (Karr et al. 1986) ………………………………………18 Table 3: List of fishes potentially occurring in aquatic habitats in and around Stones River National Battlefield………………………………………………………………..19 Table 4: Fish species list (by site) of aquatic habitats at STRI (October 2004 – August 2006). MF = McFadden’s Ford, KP = King Pond, RB = Redoubt Brannan, UP = Unnamed Pond at Redoubt Brannan, LC = Lytle Creek at Fortress Rosecrans……...….22 Table 5: Fish Species Richness estimates for the 3 riverine reaches of STRI and a composite estimate for STRI as a whole…………………………………………………24 Table 6: Index of Biotic Integrity (IBI) scores for three stream reaches at Stones River National Battlefield during August 2005………………………………………………...25 Table 7: Temperature and water chemistry of four of the STRI sample sites for each sampling date…………………………………………………………………………….26 Table 8 : Total length estimates of specific habitat types at each riverine sample site. -

C:\Fish\Eastern Sand Darter Sa.Wpd
EASTERN SAND DARTER STATUS ASSESSMENT Prepared by: David Grandmaison and Joseph Mayasich Natural Resources Research Institute University of Minnesota 5013 Miller Trunk Highway Duluth, MN 55811-1442 and David Etnier Ecology and Evolutionary Biology University of Tennessee 569 Dabney Hall Knoxville, TN 37996-1610 Prepared for: U.S. Fish and Wildlife Service Region 3 1 Federal Drive Fort Snelling, MN 55111 January 2004 NRRI Technical Report No. NRRI/TR-2003/40 DISCLAIMER This document is a compilation of biological data and a description of past, present, and likely future threats to the eastern sand darter, Ammocrypta pellucida (Agassiz). It does not represent a decision by the U.S. Fish and Wildlife Service (Service) on whether this taxon should be designated as a candidate species for listing as threatened or endangered under the Federal Endangered Species Act. That decision will be made by the Service after reviewing this document; other relevant biological and threat data not included herein; and all relevant laws, regulations, and policies. The result of the decision will be posted on the Service's Region 3 Web site (refer to: http://midwest.fws.gov/eco_serv/endangrd/lists/concern.html). If designated as a candidate species, the taxon will subsequently be added to the Service's candidate species list that is periodically published in the Federal Register and posted on the World Wide Web (refer to: http://endangered.fws.gov/wildlife.html). Even if the taxon does not warrant candidate status it should benefit from the conservation recommendations that are contained in this document. ii TABLE OF CONTENTS DISCLAIMER................................................................... -

Habitat Suitability and Detection Probability of Longnose Darter (Percina Nasuta) in Oklahoma
HABITAT SUITABILITY AND DETECTION PROBABILITY OF LONGNOSE DARTER (PERCINA NASUTA) IN OKLAHOMA By COLT TAYLOR HOLLEY Bachelor of Science in Natural Resource Ecology and Management Oklahoma State University Stillwater, OK 2016 Submitted to the Faculty of the Graduate College of the Oklahoma State University in partial fulfillment of the requirements for the Degree of MASTER OF SCIENCE December, 2018 HABITAT SUITABILITY AND DETECTION PROBABILITY OF LONGNOSE DARTER (PERCINA NASUTA) IN OKLAHOMA Thesis Approved: Dr. James M. Long Thesis Advisor Dr. Shannon Brewer Dr. Monica Papeş ii ACKNOWLEDGEMENTS I am truly thankful for the support of my advisor, Dr. Jim Long, throughout my time at Oklahoma State University. His motivation and confidence in me was invaluable. I also thank my committee members Dr. Shannon Brewer and Dr. Mona Papeş for their contributions to my education and for their comments that improved this thesis. I thank the Oklahoma Department of Wildlife Conservation (ODWC) for providing the funding for this project and the Oklahoma Cooperative Fish and Wildlife Research Unit (OKCFWRU) for their logistical support. I thank Tommy Hall, James Mier, Bill Rogers, Dick Rogers, and Mr. and Mrs. Terry Scott for allowing me to access Lee Creek from their properties. Much of my research could not have been accomplished without them. My field technicians Josh, Matt, and Erick made each field season enjoyable and I could not have done it without their help. The camaraderie of my friends and fellow graduate students made my time in Stillwater feel like home. I consider Dr. Andrew Taylor to be a mentor, fishing partner, and one of my closest friends. -

Download Download
The Implications of Rotenone Eradication on the Fish Community of Eagle Creek in Central Indiana William L. Fisher Water Resources Laboratory, University of Louisville, Louisville, Kentucky 40292 and James R. Gammon Department of Zoology, DePauw University Greencastle, Indiana 46135 Introduction The use of rotenone in the reclamation of fish populations first came into use in the United States in 1934 (Krumholz 1948). Since then, the eradication of undesirable fishes from ponds, lakes, and streams by the use of toxicants has become an accepted fisheries management practice. Forty-nine states, including Indiana, have reclaimed their waters by chemical methods, mainly since 1953 (Len- non et al. 1971). In the summer of 1966, Eagle Creek and its tributaries were surveyed prior to impoundment by the Division of Fish and Game of the Indiana Department of Natural Resources to determine the relative abundance of "undesirable" fish species, i.e., carp, Cyprinus carpio, and gizzard shad, Dorosoma cepedianum. It was concluded from the study that, "no part of the watershed of Eagle Creek Reservoir could be omitted from an eradication project" (McGinty 1966). Twelve years later, Eagle Creek was chosen as an experimental study stream in a Model Implementation Program (MIP) initiated by the U.S. Environmental Protection Agency and the Department of Agriculture. The goal of the study was to assess the effects of improved agricultural land-use practices on the water quali- ty and stream biota of Eagle Creek. A comparison of the data collected prior to the reclamation and impoundment of Eagle Creek with that gathered following yields some disturbing results and raises some serious questions about the validity of this accepted and commonly used reservoir management practice. -

FISHERIES REPORT: Warmwater Streams and Rivers
FISHERIES REPORT: Warmwater Streams and Rivers Tennessee Wildlife Resources Agency 2015 Region IV Report 16-11 FISHERIES REPORT: Warmwater Streams and Rivers FISHERIES REPORT REPORT NO. 16-11 WARMWATER STREAM FISHERIES REPORT REGION IV 2015 Prepared by Bart D. Carter Rick D. Bivens Carl E. Williams and James W. Habera Page 1 FISHERIES REPORT: Warmwater Streams and Rivers TENNESSEE WILDLIFE RESOURCES AGENCY Development of this report was financed in part by funds from Federal Aid in Fish and Wildlife Restoration (TWRA Project 4350) (Public Law 91-503) as documented in Federal Aid Project FW-6. This program receives Federal Aid in Fish and Wildlife Restoration. Under Title VI of the Civil Rights Act of 1964 and Section 504 of the Interior prohibits discrimination on the basis of race, color, national origin, or handicap. If you believe you have been discriminated against in any program, activity, or facility as described above, or if you desire further information, please write to: Office of Equal Opportunity, U.S. Department of the Interior, Washington D.C. 20240. Cover: Lake Sturgeon find a new home in the French Broad River above Douglas Reservoir. Releases were made in the French Broad River in 2014 and 2015. Page 2 FISHERIES REPORT: Warmwater Streams and Rivers TABLE OF CONTENTS Page INTRODUCTION 4 METHODS 5 Index of Biotic Integrity Surveys: South Indian Creek 9 Long Creek 14 Turkey Creek 17 Pigeon River 22 Little River 33 North Cumberland Habitat Conservation Plan Monitoring: 45 Straight Fork Jake Branch Hudson Branch Terry Creek Stinking Creek Jennings Creek Louse Creek Special Project: 53 Tennessee Dace Distribution Survey Sport Fish Survey: French Broad River 55 Holston River 67 LITERATURE CITED 84 Page 3 FISHERIES REPORT: Warmwater Streams and Rivers INTRODUCTION The fish fauna of Tennessee is the most diverse in the United States, with approximately 307 species of native fish and about 30 to 33 introduced species (Etnier and Starnes 1993). -

Stability, Persistence and Habitat Associations of the Pearl Darter Percina Aurora in the Pascagoula River System, Southeastern USA
Vol. 36: 99–109, 2018 ENDANGERED SPECIES RESEARCH Published June 13 https://doi.org/10.3354/esr00897 Endang Species Res OPENPEN ACCESSCCESS Stability, persistence and habitat associations of the pearl darter Percina aurora in the Pascagoula River System, southeastern USA Scott R. Clark1,4,*, William T. Slack2, Brian R. Kreiser1, Jacob F. Schaefer1, Mark A. Dugo3 1Department of Biological Sciences, The University of Southern Mississippi, Hattiesburg, Mississippi 39406, USA 2US Army Engineer Research and Development Center, Environmental Laboratory EEA, Vicksburg, Mississippi 39180, USA 3Mississippi Valley State University, Department of Natural Sciences and Environmental Health, Itta Bena, Mississippi 38941, USA 4Present address: Department of Biology and Museum of Southwestern Biology, University of New Mexico, Albuquerque, New Mexico 87131, USA ABSTRACT: The southeastern United States represents one of the richest collections of aquatic biodiversity worldwide; however, many of these taxa are under an increasing threat of imperil- ment, local extirpation, or extinction. The pearl darter Percina aurora is a small-bodied freshwater fish endemic to the Pearl and Pascagoula river systems of Mississippi and Louisiana (USA). The last collected specimen from the Pearl River drainage was taken in 1973, and it now appears that populations in this system are likely extirpated. This reduced the historical range of this species by approximately 50%, ultimately resulting in federal protection under the US Endangered Species Act in 2017. To better understand the current distribution and general biology of extant popula- tions, we analyzed data collected from a series of surveys conducted in the Pascagoula River drainage from 2000 to 2016. Pearl darters were captured at relatively low abundance (2.4 ± 4.0 ind. -

ECOLOGY of NORTH AMERICAN FRESHWATER FISHES
ECOLOGY of NORTH AMERICAN FRESHWATER FISHES Tables STEPHEN T. ROSS University of California Press Berkeley Los Angeles London © 2013 by The Regents of the University of California ISBN 978-0-520-24945-5 uucp-ross-book-color.indbcp-ross-book-color.indb 1 44/5/13/5/13 88:34:34 AAMM uucp-ross-book-color.indbcp-ross-book-color.indb 2 44/5/13/5/13 88:34:34 AAMM TABLE 1.1 Families Composing 95% of North American Freshwater Fish Species Ranked by the Number of Native Species Number Cumulative Family of species percent Cyprinidae 297 28 Percidae 186 45 Catostomidae 71 51 Poeciliidae 69 58 Ictaluridae 46 62 Goodeidae 45 66 Atherinopsidae 39 70 Salmonidae 38 74 Cyprinodontidae 35 77 Fundulidae 34 80 Centrarchidae 31 83 Cottidae 30 86 Petromyzontidae 21 88 Cichlidae 16 89 Clupeidae 10 90 Eleotridae 10 91 Acipenseridae 8 92 Osmeridae 6 92 Elassomatidae 6 93 Gobiidae 6 93 Amblyopsidae 6 94 Pimelodidae 6 94 Gasterosteidae 5 95 source: Compiled primarily from Mayden (1992), Nelson et al. (2004), and Miller and Norris (2005). uucp-ross-book-color.indbcp-ross-book-color.indb 3 44/5/13/5/13 88:34:34 AAMM TABLE 3.1 Biogeographic Relationships of Species from a Sample of Fishes from the Ouachita River, Arkansas, at the Confl uence with the Little Missouri River (Ross, pers. observ.) Origin/ Pre- Pleistocene Taxa distribution Source Highland Stoneroller, Campostoma spadiceum 2 Mayden 1987a; Blum et al. 2008; Cashner et al. 2010 Blacktail Shiner, Cyprinella venusta 3 Mayden 1987a Steelcolor Shiner, Cyprinella whipplei 1 Mayden 1987a Redfi n Shiner, Lythrurus umbratilis 4 Mayden 1987a Bigeye Shiner, Notropis boops 1 Wiley and Mayden 1985; Mayden 1987a Bullhead Minnow, Pimephales vigilax 4 Mayden 1987a Mountain Madtom, Noturus eleutherus 2a Mayden 1985, 1987a Creole Darter, Etheostoma collettei 2a Mayden 1985 Orangebelly Darter, Etheostoma radiosum 2a Page 1983; Mayden 1985, 1987a Speckled Darter, Etheostoma stigmaeum 3 Page 1983; Simon 1997 Redspot Darter, Etheostoma artesiae 3 Mayden 1985; Piller et al. -
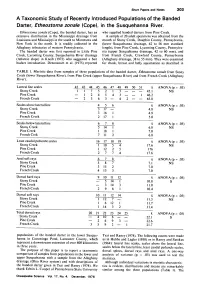
A Taxonomic Study of Recently Introduced Populations of The
Short Papers and Notes 303 A TaxonomicStudy of RecentlyIntroduced Populations of the Banded Darter,Etheostoma zonale (Cope), in the Susquehanna River. Etheostoma zonale (Cope), the banded darter, has an who supplied banded darters from Pine Creek. extensive distribution in the Mississippi drainage from A sample of 20 adult specimens was obtained from the Louisiana and Mississippi in the south to Minnesota and mouth of Stony Creek, Dauphin County, Pennsylvania, New York in the north. It is readily collected in the (lower Susquehanna drainage, 42 to 56 mm standard Allegheny tributaries of western Pennsylvania. length), from Pine Creek, Lycoming County, Pennsylva- The banded darter was first reported in Little Pine nia (upper Susquehanna drainage, 42 to 60 mm), and Creek, Lycoming County, Susquehanna River drainage from French Creek, Crawford County, Pennsylvania (Atlantic slope) in Kneib (1972) who suggested a bait (Allegheny drainage, 38 to 55 mm). They were examined bucket introduction. Denoncourt et al. (1975) reported for cheek, breast and belly squamation as described in TABLE 1. Meristic data from samples of three populations of the banded darter, Etheostoma zonale from Stony Creek (lower Susquehanna River), from Pine Creek (upper Susquehanna River) and from French Creek (Allegheny River). Lateral line scales 42 43 44 45 46 47 48 49 50 51 k ANOVA (p = .05) Stony Creek 1 1 7 5 2 1 3 - -- 45.1 NS Pine Creek - 1 2 4 5 4 3 - - 1 46.2 French Creek - 2 3 6 3 - 4 2 - - 45.8 Scales above lateral line 4 5 6 R ANOVA (p = .05) Stony Creek 3 -

Spatial Criteria Used in IUCN Assessment Overestimate Area of Occupancy for Freshwater Taxa
Spatial Criteria Used in IUCN Assessment Overestimate Area of Occupancy for Freshwater Taxa By Jun Cheng A thesis submitted in conformity with the requirements for the degree of Masters of Science Ecology and Evolutionary Biology University of Toronto © Copyright Jun Cheng 2013 Spatial Criteria Used in IUCN Assessment Overestimate Area of Occupancy for Freshwater Taxa Jun Cheng Masters of Science Ecology and Evolutionary Biology University of Toronto 2013 Abstract Area of Occupancy (AO) is a frequently used indicator to assess and inform designation of conservation status to wildlife species by the International Union for Conservation of Nature (IUCN). The applicability of the current grid-based AO measurement on freshwater organisms has been questioned due to the restricted dimensionality of freshwater habitats. I investigated the extent to which AO influenced conservation status for freshwater taxa at a national level in Canada. I then used distribution data of 20 imperiled freshwater fish species of southwestern Ontario to (1) demonstrate biases produced by grid-based AO and (2) develop a biologically relevant AO index. My results showed grid-based AOs were sensitive to spatial scale, grid cell positioning, and number of records, and were subject to inconsistent decision making. Use of the biologically relevant AO changed conservation status for four freshwater fish species and may have important implications on the subsequent conservation practices. ii Acknowledgments I would like to thank many people who have supported and helped me with the production of this Master’s thesis. First is to my supervisor, Dr. Donald Jackson, who was the person that inspired me to study aquatic ecology and conservation biology in the first place, despite my background in environmental toxicology. -

Reproductive Ecology and Habitat Preference of the Leopard Darter, Percina Pantherina
REPRODUCTIVE ECOLOGY AND HABITAT PREFERENCE OF THE LEOPARD DARTER, PERCINA PANTHERINA By PAUL WILLIAM /~AMES Bachelor of Science University of Kansas Lawrence, Kansas 1981 ·4::er of Science ...1.issouri State University 3pringfield, Missouri 1983 Submitted to the Faculty of the Graduate College of the Oklahoma State University in partial fulfillment of the requirements for the·Degree of DOCTOR OF PHILOSOPHY July, 1989 . - ~· ,• ) "' Oklahoma State Univ. Lihra1 REPRODUCTIVE ECOLOGY AND HABITAT PREFERENCE OF THE LEOPARD DARTER, PERCINA PANTHERINA Thesis Approved: ii ACKNOWLEDGMENTS I wish to thank my advisor, Dr. o. Eugene Maughan, for giving me the opportunity to work on this project and for his encouragement throughout my graduate program. I would also like to thank the members of my graduate committee, Dr. William A. Drew, Dr. Anthony A. Echelle, Dr. Rudolph J. Miller, and Dr. Alexander v. Zale, for their professional and personal advice throughout the course of the study. I wish to extend my sincere gratitude to the u. s. Fish and Wildlife Service, the Oklahoma Department of Wildlife Conservation, and the Oklahoma Cooperative Fish and Wildlife Research Unit for providing financial and technical support for the study. I am especially grateful to Mr. Frank James of the Oklahoma Department of Wildlife Conservation's McCurtain County Wilderness Area for his friendship and hospitality during extended field trips. A sincere thanks goes to Rick Horton, Steve O'Donnell, and Todd Phillips for their help in the field and laboratory. A special thanks goes to Stuart Leon for helping with the development of many of the field and data analysis techniques used in this study.