Biostratigraphy, Lithostratigraphy, Ammonite Taxonomy and Microfacies Analysis of the Middle and Upper Jurassic of Norteastern Iran
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Cambrian Phytoplankton of the Brunovistulicum – Taxonomy and Biostratigraphy
MONIKA JACHOWICZ-ZDANOWSKA Cambrian phytoplankton of the Brunovistulicum – taxonomy and biostratigraphy Polish Geological Institute Special Papers,28 WARSZAWA 2013 CONTENTS Introduction...........................................................6 Geological setting and lithostratigraphy.............................................8 Summary of Cambrian chronostratigraphy and acritarch biostratigraphy ...........................13 Review of previous palynological studies ...........................................17 Applied techniques and material studied............................................18 Biostratigraphy ........................................................23 BAMA I – Pulvinosphaeridium antiquum–Pseudotasmanites Assemblage Zone ....................25 BAMA II – Asteridium tornatum–Comasphaeridium velvetum Assemblage Zone ...................27 BAMA III – Ichnosphaera flexuosa–Comasphaeridium molliculum Assemblage Zone – Acme Zone .........30 BAMA IV – Skiagia–Eklundia campanula Assemblage Zone ..............................39 BAMA V – Skiagia–Eklundia varia Assemblage Zone .................................39 BAMA VI – Volkovia dentifera–Liepaina plana Assemblage Zone (Moczyd³owska, 1991) ..............40 BAMA VII – Ammonidium bellulum–Ammonidium notatum Assemblage Zone ....................40 BAMA VIII – Turrisphaeridium semireticulatum Assemblage Zone – Acme Zone...................41 BAMA IX – Adara alea–Multiplicisphaeridium llynense Assemblage Zone – Acme Zone...............42 Regional significance of the biostratigraphic -

Post-Carboniferous Stratigraphy, Northeastern Alaska by R
Post-Carboniferous Stratigraphy, Northeastern Alaska By R. L. DETTERMAN, H. N. REISER, W. P. BROSGE,and]. T. DUTRO,JR. GEOLOGICAL SURVEY PROFESSIONAL PAPER 886 Sedirnentary rocks of Permian to Quaternary age are named, described, and correlated with standard stratigraphic sequences UNITED STATES GOVERNMENT PRINTING OFFICE, WASHINGTON 1975 UNITED STATES DEPARTMENT OF THE INTERIOR ROGERS C. B. MORTON, Secretary GEOLOGICAL SURVEY V. E. McKelvey, Director Library of Congress Cataloging in Publication Data Detterman, Robert L. Post-Carboniferous stratigraphy, northeastern Alaska. (Geological Survey Professional Paper 886) Bibliography: p. 45-46. Supt. of Docs. No.: I 19.16:886 1. Geology-Alaska. I. Detterman, Robert L. II. Series: United States. Geological Survey. Professional Paper 886. QE84.N74P67 551.7'6'09798 74-28084 For sale by the Superintendent of Documents, U.S. Government Printing Office Washington, D.C. 20402 Stock Number 024-001-02687-2 CONTENTS Page Page Abstract __ _ _ _ _ __ __ _ _ _ _ _ _ _ _ _ _ _ _ __ __ _ _ _ _ _ _ __ __ _ _ __ __ __ _ _ _ _ __ 1 Stratigraphy__:_Continued Introduction __________ ----------____ ----------------____ __ 1 Kingak Shale ---------------------------------------- 18 Purpose and scope ----------------------~------------- 1 Ignek Formation (abandoned) -------------------------- 20 Geographic setting ------------------------------------ 1 Okpikruak Formation (geographically restricted) ________ 21 Previous work and acknowledgments ------------------ 1 Kongakut Formation ---------------------------------- -

New and Poorly Known Perisphinctoidea (Ammonitina) from the Upper Tithonian of Le Chouet (Drôme, SE France)
Volumina Jurassica, 2014, Xii (1): 113–128 New and poorly known Perisphinctoidea (Ammonitina) from the Upper Tithonian of Le Chouet (Drôme, SE France) Luc G. BULOT1, Camille FRAU2, William A.P. WIMBLEDON3 Key words: Ammonoidea, Ataxioceratidae, Himalayitidae, Neocomitidae, Upper Tithonian, Le Chouet, South-East France. Abstract. The aim of this paper is to document the ammonite fauna of the upper part of the Late Tithonian collected at the key section of Le Chouet (Drôme, SE France). Emphasis is laid on new and poorly known Ataxioceratidae, Himalayitidae and Neocomitidae from the upper part of the Tithonian. Among the Ataxioceratidae, a new account on the taxonomy and relationship between Paraulacosphinctes Schindewolf and Moravisphinctes Tavera is presented. Regarding the Himalayitidae, the range and content of Micracanthoceras Spath is discussed and two new genera are introduced: Ardesciella gen. nov., for a group of Mediterranean ammonites that is homoeomorphic with the Andean genus Corongoceras Spath, and Pratumidiscus gen. nov. for a specimen that shows morphological similarities with the Boreal genera Riasanites Spath and Riasanella Mitta. Finally, the occurrence of Neocomitidae in the uppermost Tithonian is documented by the presence of the reputedly Berriasian genera Busnardoiceras Tavera and Pseudargentiniceras Spath. INTRODUCTION known Perisphinctoidea from the Upper Tithonian of this reference section. Additional data on the Himalayitidae in- The unique character of the ammonite fauna of Le Chouet cluding the description and discussion of Boughdiriella (near Les Près, Drôme, France) (Fig. 1) has already been chouetensis gen. nov. sp. nov. are to be published elsewhere outlined by Le Hégarat (1973), but, so far, only a handful of (Frau et al., 2014). -

Cause and Consequence of Recurrent Early Jurassic Anoxia Following The
[Palaeontology, Vol. 56, Part 4, 2013, pp. 685–709] MICROBES, MUD AND METHANE: CAUSE AND CONSEQUENCE OF RECURRENT EARLY JURASSIC ANOXIA FOLLOWING THE END-TRIASSIC MASS EXTINCTION by BAS VAN DE SCHOOTBRUGGE1*, AVIV BACHAN2, GUILLAUME SUAN3, SYLVAIN RICHOZ4 and JONATHAN L. PAYNE2 1Palaeo-environmental Dynamics Group, Institute of Geosciences, Goethe University Frankfurt, Altenhofer€ Allee 1, 60438, Frankfurt am Main, Germany; email: [email protected] 2Geological and Environmental Sciences, Stanford University, 450 Serra Mall, Stanford, CA 94305, USA; emails: [email protected], [email protected] 3UMR, CNRS 5276, LGLTPE, Universite Lyon 1, F-69622, Villeurbanne, France; email: [email protected] 4Academy of Sciences, University of Graz, Heinrichstraße 26, 8020, Graz, Austria; email: [email protected] *Corresponding author. Typescript received 19 January 2012; accepted in revised form 23 January 2013 Abstract: The end-Triassic mass extinction (c. 201.6 Ma) Toarcian events are marked by important changes in phyto- was one of the five largest mass-extinction events in the his- plankton assemblages from chromophyte- to chlorophyte- tory of animal life. It was also associated with a dramatic, dominated assemblages within the European Epicontinental long-lasting change in sedimentation style along the margins Seaway. Phytoplankton changes occurred in association with of the Tethys Ocean, from generally organic-matter-poor the establishment of photic-zone euxinia, driven by a general sediments during the -

Lower Jurassic to Lower Middle Jurassic Succession at Kopy Sołtysie and Płaczliwa Skała in the Eastern Tatra Mts (Western
Volumina Jurassica, 2013, Xi: 19–58 Lower Jurassic to lower Middle Jurassic succession at Kopy Sołtysie and Płaczliwa Skała in the eastern Tatra Mts (Western Carpathians) of Poland and Slovakia: stratigraphy, facies and ammonites Jolanta IWAŃCZUK1, Andrzej IWANOW1, Andrzej WIERZBOWSKI1 Key words: stratigraphy, Lower to Middle Jurassic, ammonites, microfacies, correlations, Tatra Mts, Western Carpathians. Abstract. The Lower Jurassic and the lower part of the Middle Jurassic deposits corresponding to the Sołtysia Marlstone Formation of the Lower Subtatric (Krížna) nappe in the Kopy Sołtysie mountain range of the High Tatra Mts and the Płaczliwa Skała (= Ždziarska Vidla) mountain of the Belianske Tatra Mts in the eastern part of the Tatra Mts in Poland and Slovakia are described. The work concentrates both on their lithological and facies development as well as their ammonite faunal content and their chronostratigraphy. These are basinal de- posits which show the dominant facies of the fleckenkalk-fleckenmergel type and reveal the succession of several palaeontological microfacies types from the spiculite microfacies (Sinemurian–Lower Pliensbachian, but locally also in the Bajocian), up to the radiolarian microfacies (Upper Pliensbachian and Toarcian, Bajocian–Bathonian), and locally the Bositra (filament) microfacies (Bajocian– Bathonian). In addition, there appear intercalations of detrital deposits – both bioclastic limestones and breccias – formed by downslope transport from elevated areas (junction of the Sinemurian and Pliensbachian, Upper Toarcian, and Bajocian). The uppermost Toarcian – lowermost Bajocian interval is represented by marly-shaly deposits with a marked admixture of siliciclastic material. The deposits are correlated with the coeval deposits of the Lower Subtatric nappe of the western part of the Tatra Mts (the Bobrowiec unit), as well as with the autochthonous-parachthonous Hightatric units, but also with those of the Czorsztyn and Niedzica successions of the Pieniny Klippen Belt, in Poland. -

Appendix 3.Pdf
A Geoconservation perspective on the trace fossil record associated with the end – Ordovician mass extinction and glaciation in the Welsh Basin Item Type Thesis or dissertation Authors Nicholls, Keith H. Citation Nicholls, K. (2019). A Geoconservation perspective on the trace fossil record associated with the end – Ordovician mass extinction and glaciation in the Welsh Basin. (Doctoral dissertation). University of Chester, United Kingdom. Publisher University of Chester Rights Attribution-NonCommercial-NoDerivatives 4.0 International Download date 26/09/2021 02:37:15 Item License http://creativecommons.org/licenses/by-nc-nd/4.0/ Link to Item http://hdl.handle.net/10034/622234 International Chronostratigraphic Chart v2013/01 Erathem / Era System / Period Quaternary Neogene C e n o z o i c Paleogene Cretaceous M e s o z o i c Jurassic M e s o z o i c Jurassic Triassic Permian Carboniferous P a l Devonian e o z o i c P a l Devonian e o z o i c Silurian Ordovician s a n u a F y r Cambrian a n o i t u l o v E s ' i k s w o Ichnogeneric Diversity k p e 0 10 20 30 40 50 60 70 S 1 3 5 7 9 11 13 15 17 19 21 n 23 r e 25 d 27 o 29 M 31 33 35 37 39 T 41 43 i 45 47 m 49 e 51 53 55 57 59 61 63 65 67 69 71 73 75 77 79 81 83 85 87 89 91 93 Number of Ichnogenera (Treatise Part W) Ichnogeneric Diversity 0 10 20 30 40 50 60 70 1 3 5 7 9 11 13 15 17 19 21 n 23 r e 25 d 27 o 29 M 31 33 35 37 39 T 41 43 i 45 47 m 49 e 51 53 55 57 59 61 c i o 63 z 65 o e 67 a l 69 a 71 P 73 75 77 79 81 83 n 85 a i r 87 b 89 m 91 a 93 C Number of Ichnogenera (Treatise Part W) -
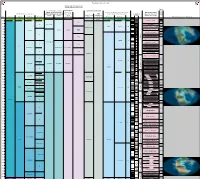
Expanded Jurassic Timescale
TimeScale Creator 2012 chart Russian and Ural regional units Russia Platform regional units Calca Jur-Cret boundary regional Russia Platform East Asian regional units reous stages - British and Boreal Stages (Jur- Australia and New Zealand regional units Marine Macrofossils Nann Standard Chronostratigraphy British regional Boreal regional Cret, Perm- Japan New Zealand Chronostratigraphy Geomagnetic (Mesozoic-Paleozoic) ofossil stages stages Carb & South China (Neogene & Polarity Tethyan Ammonoids s Ma Period Epoch Age/Stage Substage Cambrian) stages Cret) NZ Series NZ Stages Global Reconstructions (R. Blakey) Ryazanian Ryazanian Ryazanian [ no stages M17 CC2 Cretaceous Early Berriasian E Kochian Taitai Um designated ] M18 CC1 145 Berriasella jacobi M19 NJT1 Late M20 7b 146 Lt Portlandian M21 Durangites NJT1 M22 7a 147 Oteke Puaroan Op M22A Micracanthoceras microcanthum NJT1 Penglaizhenian M23 6b 148 Micracanthoceras ponti / Volgian Volgian Middle M24 Burckhardticeras peroni NJT1 Tithonian M24A 6a 149 M24B Semiformiceras fallauxi NJT15 M25 b E 150 M25A Semiformiceras semiforme NJT1 5a Early M26 Semiformiceras darwini 151 lt-Oxf N M-Sequence Hybonoticeras hybonotum 152 Ohauan Ko lt-Oxf R Kimmeridgian Hybonoticeras beckeri 153 m- Lt Late Oxf N Aulacostephanus eudoxus 154 m- NJT14 Late Aspidoceras acanthicum Oxf R Kimmeridgian Kimmeridgian Kimmeridgian Crussoliceras divisum 155 155.431 Card- N Ataxioceras hypselocyclum 156 E Early e-Oxf Sutneria platynota R Idoceras planula Suiningian 157 Cal- Oxf N Epipeltoceras bimammatum 158 lt- Lt Callo -

A Review of the Classification of Jurassic Aspidoceratid Ammonites – the Superfamily Aspidoceratoidea
VOLUMINA JURASSICA, 2020, XVIII (1): 47–52 DOI: 10.7306/VJ.18.4 A review of the classification of Jurassic aspidoceratid ammonites – the Superfamily Aspidoceratoidea Horacio PARENT1, Günter SCHWEIGERT2, Armin SCHERZINGER3 Key words: Superfamily Aspidoceratoidea, Aspidoceratidae, Epipeltoceratinae emended, Peltoceratidae, Gregoryceratinae nov. subfam. Abstract. The aspidoceratid ammonites have been traditionally included in the superfamily Perisphinctoidea. However, the basis of this is unclear for they bear unique combinations of characters unknown in typical perisphinctoids: (1) the distinct laevaptychus, (2) stout shells with high growth rate of the whorl section area, (3) prominent ornamentation with tubercles, spines and strong growth lines running in parallel over strong ribs, (4) lack of constrictions, (5) short to very short bodychamber, and (6) sexual dimorphism characterized by minia- turized microconchs and small-sized macroconchs besides the larger ones, including changes of sex during ontogeny in many cases. Considering the uniqueness of these characters we propose herein to raise the family Aspidoceratidae to the rank of a superfamily Aspi- doceratoidea, ranging from the earliest Late Callovian to the Early Berriasian Jacobi Zone. The new superfamily includes two families, Aspidoceratidae (Aspidoceratinae, Euaspidoceratinae, Epipeltoceratinae and Hybonoticeratinae), and Peltoceratidae (Peltoceratinae and Gregoryceratinae nov. subfam.). The highly differentiated features of the aspidoceratoids indicate that their life-histories -

First Record of Non-Mineralized Cephalopod Jaws and Arm Hooks
Klug et al. Swiss J Palaeontol (2020) 139:9 https://doi.org/10.1186/s13358-020-00210-y Swiss Journal of Palaeontology RESEARCH ARTICLE Open Access First record of non-mineralized cephalopod jaws and arm hooks from the latest Cretaceous of Eurytania, Greece Christian Klug1* , Donald Davesne2,3, Dirk Fuchs4 and Thodoris Argyriou5 Abstract Due to the lower fossilization potential of chitin, non-mineralized cephalopod jaws and arm hooks are much more rarely preserved as fossils than the calcitic lower jaws of ammonites or the calcitized jaw apparatuses of nautilids. Here, we report such non-mineralized fossil jaws and arm hooks from pelagic marly limestones of continental Greece. Two of the specimens lie on the same slab and are assigned to the Ammonitina; they represent upper jaws of the aptychus type, which is corroborated by fnds of aptychi. Additionally, one intermediate type and one anaptychus type are documented here. The morphology of all ammonite jaws suggest a desmoceratoid afnity. The other jaws are identifed as coleoid jaws. They share the overall U-shape and proportions of the outer and inner lamellae with Jurassic lower jaws of Trachyteuthis (Teudopseina). We also document the frst belemnoid arm hooks from the Tethyan Maastrichtian. The fossils described here document the presence of a typical Mesozoic cephalopod assemblage until the end of the Cretaceous in the eastern Tethys. Keywords: Cephalopoda, Ammonoidea, Desmoceratoidea, Coleoidea, Maastrichtian, Taphonomy Introduction as jaws, arm hooks, and radulae are occasionally found Fossil cephalopods are mainly known from preserved (Matern 1931; Mapes 1987; Fuchs 2006a; Landman et al. mineralized parts such as aragonitic phragmocones 2010; Kruta et al. -

301083243.Pdf
AMEGHINIANA ISSN 0002-7014 Revista de la Asociación Paleontológica Argentina Tomo XII Diciembre de 1975 N?4 THE INDO-PACIFIC AMMONITE MAY AITES IN THE OXFORDIAN OF THESOUTHERN ANDES By P. N. STIPANICIC', G. E. G. WESTERMANN 2 and A. C. RJCCARDI3 ABSTRACT: Oxfordian Iitho- and biostratigraphy of the Chilean and Argentine Andes is reviewed (P. N. Stipanicic). Within the Chacay Group, the Lower to basal Upper Oxfordian La Manga Formation, below, mostly detrital and biogenic, and the Upper Oxfordian Au- quilco Formation, above, mainly chemical, are distinguished. The La Manga Formation (with Gryphaea calceola lumachelle) is rich in ammonite faunas, particularly of thc upper Cordatum to lower Canaliculatum Zones. In Neuquén and Mendoza provinces of Argentina, the Pli- catilis Zone or Middle Oxfordian has yielded Perísphinctes spp., Euaspidoceras spp., Aspido- ceras spp., together with Mayaítes (Araucanites ) stípanícfcí, M. (A.) reyesi, and M. (A.) mulai, Westermann et Riccardi subgen. et spp. nov. The first find of Mayaitidae outside the Indo-Pacific province is discussed in light of _plate-tectonic theory. RESUMEN: La revisron Iito- y bioestratigráfica del Oxfordiano de los Andes de Argentina y Chile (P. N. Stipanicic) ha permitido reconocer dentro del Grupo Chacay: 1) abajo, la Formación La Manga, mayormente detrítica y biogénica, del Oxfordiano inferior-superior basa!, y 2) arriba, la Formación Auquilco, mayormente química, del Oxfordiano superior. La For- mación La Manga (con lumachelas de Gryphaea calceola) contiene abundante cantidad de amonitas, particularmente de las Zonas de Cordatum superior a Canaliculatum inferior. En las provincias de Mendoza y Neuquén,Argentina, la Zona de Plicatilis (Oxfordiano medio) contiene Perispbinctes spp., Euaspidoceras spp., Aspidoceras spp., conjuntamente con Mayaites (Araucanites) stipanicici, M. -

Revisión De Los Ammonoideos Del Lías Español Depositados En El Museo Geominero (ITGE, Madrid)
Boletín Geológico y Minero. Vol. 107-2 Año 1996 (103-124) El Instituto Tecnológico Geominero de España hace presente que las opiniones y hechos con signados en sus publicaciones son de la exclusi GEOLOGIA va responsabilidad de los autores de los trabajos. Revisión de los Ammonoideos del Lías español depositados en el Museo Geominero (ITGE, Madrid). Por J. BERNAD (*) y G. MARTINEZ. (**) RESUMEN Se revisan desde el punto de vista taxonómico, los fósiles de ammonoideos correspondientes al Lías español que se encuentran depositados en el Museo Geominero. La colección está compuesta por ejemplares procedentes de 67 localida des españolas, pertenecientes a colecciones de diferentes autores. Se identifican los ordenes Phylloceratina, Lytoceratina y Ammonitina, las familias Phylloceratidae, Echioceratidae, eoderoceratidae, Liparoceratidae, Amaltheidae, Dactyliocerati Los derechos de propiedad de los trabajos dae, Hildoceratidae y Hammatoceratidae y las subfamilias Xipheroceratinae, Arieticeratinae, Harpoceratinae, Hildocerati publicados en esta obra fueron cedidos por nae, Grammoceratinae, Phymatoceratinae y Hammatoceratinae correspondientes a los pisos Sinemuriense, Pliensbachien los autores al Instituto Tecnológico Geomi se y Toarciense. nero de España Oueda hecho el depósito que marca la ley. Palabras clave: Ammonoidea, Taxonomía, Lías, España, Museo Geominero. ABSTRACT The Spanish Liassic ammonoidea fossil collections of the Geominero Museum is revised under a taxonomic point of view. The collection includes specimens from 67 Spanish -

Palaeoecology and Palaeoenvironments of the Middle Jurassic to Lowermost Cretaceous Agardhfjellet Formation (Bathonian–Ryazanian), Spitsbergen, Svalbard
NORWEGIAN JOURNAL OF GEOLOGY Vol 99 Nr. 1 https://dx.doi.org/10.17850/njg99-1-02 Palaeoecology and palaeoenvironments of the Middle Jurassic to lowermost Cretaceous Agardhfjellet Formation (Bathonian–Ryazanian), Spitsbergen, Svalbard Maayke J. Koevoets1, Øyvind Hammer1 & Crispin T.S. Little2 1Natural History Museum, University of Oslo, P.O. Box 1172 Blindern, 0318 Oslo, Norway. 2School of Earth and Environment, University of Leeds, Leeds LS2 9JT, United Kingdom. E-mail corresponding author (Maayke J. Koevoets): [email protected] We describe the invertebrate assemblages in the Middle Jurassic to lowermost Cretaceous of the Agardhfjellet Formation present in the DH2 rock-core material of Central Spitsbergen (Svalbard). Previous studies of the Agardhfjellet Formation do not accurately reflect the distribution of invertebrates throughout the unit as they were limited to sampling discontinuous intervals at outcrop. The rock-core material shows the benthic bivalve fauna to reflect dysoxic, but not anoxic environments for the Oxfordian–Lower Kimmeridgian interval with sporadic monospecific assemblages of epifaunal bivalves, and more favourable conditions in the Volgian, with major increases in abundance and diversity of Hartwellia sp. assemblages. Overall, the new information from cores shows that abundance, diversity and stratigraphic continuity of the fossil record in the Upper Jurassic of Spitsbergen are considerably higher than indicated in outcrop studies. The inferred life positions and feeding habits of the benthic fauna refine our understanding of the depositional environments of the Agardhfjellet Formation. The pattern of occurrence of the bivalve genera is correlated with published studies of Arctic localities in East Greenland and northern Siberia and shows similarities in palaeoecology with the former but not the latter.