Appendix 3.Pdf
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

On the Continuity of Background and Mass Extinction
Paleobiology, 29(4), 2003, pp. 455±467 On the continuity of background and mass extinction Steve C. Wang Abstract.ÐDo mass extinctions grade continuously into the background extinctions occurring throughout the history of life, or are they a fundamentally distinct phenomenon that cannot be explained by processes responsible for background extinction? Various criteria have been proposed for addressing this question, including approaches based on physical mechanisms, ecological se- lectivity, and statistical characterizations of extinction intensities. Here I propose a framework de®ning three types of continuity of mass and background extinc- tionsÐcontinuity of cause, continuity of effect, and continuity of magnitude. I test the third type of continuity with a statistical method based on kernel density estimation. Previous statistical ap- proaches typically have examined quantitative characteristics of mass extinctions (such as metrics of extinction intensity) and compared them with the distribution of such characteristics associated with background extinctions. If mass extinctions are outliers, or are separated by a gap from back- ground extinctions, the distinctness of mass extinctions is supported. In this paper I apply Silverman's Critical Bandwidth Test to test for the continuity of mass ex- tinctions by applying kernel density estimation and bootstrap modality testing. The method im- proves on existing work based on searching for gaps in histograms, in that it does not depend on arbitrary choices of parameters (such as bin widths for histograms), and provides a direct estimate of the signi®cance of continuities or gaps in observed extinction intensities. I am thus able to test rigorously whether differences between mass extinctions and background extinctions are statisti- cally signi®cant. -
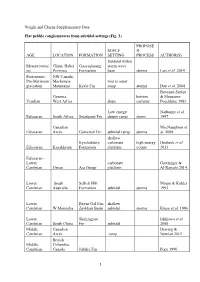
Wright and Cherns Supplementary Data Flat
Wright and Cherns Supplementary Data Flat pebble conglomerates from subtidal settings (Fig. 2) PROPOSE SHELF D AGE LOCATION FORMATION SETTING PROCESS AUTHOR(S) Subtidal within Mesoproteroz China, Hebei Gaoyuzhuang storm wave oic Province Formation base storms Luo et al. 2014 Proterozoic, NW Canada, Pre-Marinoan Mackenzie mid to outer glaciation Mountains Keele Fm ramp storms Day et al. 2004 Bertrand-Sarfati Gourma, bottom & Moussine- Vendian West Africa slope currents' Pouchkine 1983 Low energy Narbonne et al. Ediacaran South Africa Swartpunt Fm deeper ramp storm 1997 Canadian MacNaughton et Ediacaran Arctic Gametrail Fm subtidal ramp storms al. 2008 shallow Kyrshabakty carbonate high energy Heubeck et al. Ediacaran Kazakhstan Formation platform events 2013 Ediacaran - Lower carbonate Grotzinger & Cambrian Oman Ara Group platform Al-Rawahi 2014 Lower South Sellick Hill Mount & Kidder Cambrian Australia Formation subtidal storms 1993 Lower Bayan Gol Fm, shallow Cambrian W Mongolia Zavkhan Basin subtidal storms Kruse et al. 1996 Lower Shuijingtuo Ishikawa et al. Cambrian South China Fm subtidal 2008 Middle Canadian Dewing & Cambrian Arctic ramp Nowlan 2012 British Middle Columbia, Cambrian Canada Jubilee Fm Pope 1990 1 Middle Pratt & Cambrian Argentina La Laja Fm subtidal shelf tsunamis Bordonaro 2007 low energy Middle shallow Cambrian Australia Ranken Lst subtidal storms Kruse 1996 Middle Wyoming, Upper Gros Cambrian USA Ventre Shale Csonka 2009 middle upper Middle W Utah, upper Wheeler, carbonate belt - Cambrian USA Marjum fms subtidal shelf Robison 1964 Middle- Upper Supratidal to Cambrian NW China subtidal fpc storms Liang et al. 1993 Ust’- Brus,Labaz, Middle Orakta, Cambrian - Kulyumbe, Lower Ujgur and Iltyk Kouchinsky et Ordovician Siberia fms al. -

New Permian Fauna from Tropical Gondwana
ARTICLE Received 18 Jun 2015 | Accepted 18 Sep 2015 | Published 5 Nov 2015 DOI: 10.1038/ncomms9676 OPEN New Permian fauna from tropical Gondwana Juan C. Cisneros1,2, Claudia Marsicano3, Kenneth D. Angielczyk4, Roger M. H. Smith5,6, Martha Richter7, Jo¨rg Fro¨bisch8,9, Christian F. Kammerer8 & Rudyard W. Sadleir4,10 Terrestrial vertebrates are first known to colonize high-latitude regions during the middle Permian (Guadalupian) about 270 million years ago, following the Pennsylvanian Gondwanan continental glaciation. However, despite over 150 years of study in these areas, the bio- geographic origins of these rich communities of land-dwelling vertebrates remain obscure. Here we report on a new early Permian continental tetrapod fauna from South America in tropical Western Gondwana that sheds new light on patterns of tetrapod distribution. Northeastern Brazil hosted an extensive lacustrine system inhabited by a unique community of temnospondyl amphibians and reptiles that considerably expand the known temporal and geographic ranges of key subgroups. Our findings demonstrate that tetrapod groups common in later Permian and Triassic temperate communities were already present in tropical Gondwana by the early Permian (Cisuralian). This new fauna constitutes a new biogeographic province with North American affinities and clearly demonstrates that tetrapod dispersal into Gondwana was already underway at the beginning of the Permian. 1 Centro de Cieˆncias da Natureza, Universidade Federal do Piauı´, 64049-550 Teresina, Brazil. 2 Programa de Po´s-Graduac¸a˜o em Geocieˆncias, Departamento de Geologia, Universidade Federal de Pernambuco, 50740-533 Recife, Brazil. 3 Departamento de Cs. Geologicas, FCEN, Universidad de Buenos Aires, IDEAN- CONICET, C1428EHA Ciudad Auto´noma de Buenos Aires, Argentina. -

A New Species of Tiaracrinus from the Latest Emsian of Morocco and Its Phylogeny
A new species of Tiaracrinus from the latest Emsian of Morocco and its phylogeny CHRISTIAN KLUG, KENNETH DE BAETS, CAROLE JUNE NAGLIK, and JOHNNY WATERS Klug, C., De Baets, K., Naglik, C.J., and Waters, J. 2014. A new species of Tiaracrinus from the latest Emsian of Morocco and its phylogeny. Acta Palaeontologica Polonica 59 (1): 135–145. We describe a new species of the unusual crinoid Tiaracrinus, T. jeanlemenni sp. nov. from the latest Emsian of the fa- mous mudmound locality Hamar Laghdad, Morocco. It differs from the previously known species in the higher number of ribs and the vaulted rib-fields, which is corroborated by the comparison of simple quantitative characters and ratios as well as by the results of a cluster analysis and a Principal Component Analysis. Based on the new material and the published specimens, we discuss the phylogeny of the genus and suggest that T. oehlerti and T. moravicus represent the ancestral forms of this small clade. Key words: Crinoidea, mudmounds, phylogeny, morphometry, symmetry, Devonian, Morocco. Christian Klug [[email protected]] and Carole J. Naglik [[email protected]], Paläontologisches Institut und Museum, Universität Zürich, Karl Schmid-Strasse 4, CH-8006 Zürich, Switzerland; Kenneth De Baets [[email protected]], GeoZentrum Nordbayern, Fachgruppe PaläoUmwelt, Universität Erlan- gen, Loewenichstr. 28, D-91054 Erlangen, Germany; Johnny Waters [[email protected]], Department of Geology, Appalachian State University, ASU Box 32067, Boone, NC 28608-2067, USA. Received 30 November 2011, accepted 4 June 2012, available online 6 June 2012. Copyright © 2014 C. Klug et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. -

Lower Jurassic to Lower Middle Jurassic Succession at Kopy Sołtysie and Płaczliwa Skała in the Eastern Tatra Mts (Western
Volumina Jurassica, 2013, Xi: 19–58 Lower Jurassic to lower Middle Jurassic succession at Kopy Sołtysie and Płaczliwa Skała in the eastern Tatra Mts (Western Carpathians) of Poland and Slovakia: stratigraphy, facies and ammonites Jolanta IWAŃCZUK1, Andrzej IWANOW1, Andrzej WIERZBOWSKI1 Key words: stratigraphy, Lower to Middle Jurassic, ammonites, microfacies, correlations, Tatra Mts, Western Carpathians. Abstract. The Lower Jurassic and the lower part of the Middle Jurassic deposits corresponding to the Sołtysia Marlstone Formation of the Lower Subtatric (Krížna) nappe in the Kopy Sołtysie mountain range of the High Tatra Mts and the Płaczliwa Skała (= Ždziarska Vidla) mountain of the Belianske Tatra Mts in the eastern part of the Tatra Mts in Poland and Slovakia are described. The work concentrates both on their lithological and facies development as well as their ammonite faunal content and their chronostratigraphy. These are basinal de- posits which show the dominant facies of the fleckenkalk-fleckenmergel type and reveal the succession of several palaeontological microfacies types from the spiculite microfacies (Sinemurian–Lower Pliensbachian, but locally also in the Bajocian), up to the radiolarian microfacies (Upper Pliensbachian and Toarcian, Bajocian–Bathonian), and locally the Bositra (filament) microfacies (Bajocian– Bathonian). In addition, there appear intercalations of detrital deposits – both bioclastic limestones and breccias – formed by downslope transport from elevated areas (junction of the Sinemurian and Pliensbachian, Upper Toarcian, and Bajocian). The uppermost Toarcian – lowermost Bajocian interval is represented by marly-shaly deposits with a marked admixture of siliciclastic material. The deposits are correlated with the coeval deposits of the Lower Subtatric nappe of the western part of the Tatra Mts (the Bobrowiec unit), as well as with the autochthonous-parachthonous Hightatric units, but also with those of the Czorsztyn and Niedzica successions of the Pieniny Klippen Belt, in Poland. -

Climatic Shifts Drove Major Contractions in Avian Latitudinal Distributions Throughout the Cenozoic
Climatic shifts drove major contractions in avian latitudinal distributions throughout the Cenozoic Erin E. Saupea,1,2, Alexander Farnsworthb, Daniel J. Luntb, Navjit Sagooc, Karen V. Phamd, and Daniel J. Fielde,1,2 aDepartment of Earth Sciences, University of Oxford, OX1 3AN Oxford, United Kingdom; bSchool of Geographical Sciences, University of Bristol, Clifton, BS8 1SS Bristol, United Kingdom; cDepartment of Meteorology, Stockholm University, 106 91 Stockholm, Sweden; dDivision of Geological and Planetary Sciences, Caltech, Pasadena, CA 91125; and eDepartment of Earth Sciences, University of Cambridge, CB2 3EQ Cambridge, United Kingdom Edited by Nils Chr. Stenseth, University of Oslo, Oslo, Norway, and approved May 7, 2019 (received for review March 8, 2019) Many higher level avian clades are restricted to Earth’s lower lati- order avian historical biogeography invariably recover strong evi- tudes, leading to historical biogeographic reconstructions favoring a dence for an origin of most modern diversity on southern land- Gondwanan origin of crown birds and numerous deep subclades. masses (2, 6, 11). However, several such “tropical-restricted” clades (TRCs) are repre- The crown bird fossil record has unique potential to reveal sented by stem-lineage fossils well outside the ranges of their clos- where different groups of birds were formerly distributed in deep est living relatives, often on northern continents. To assess the time. Fossil evidence, for example, has long indicated that total- drivers of these geographic disjunctions, we combined ecological group representatives of clades restricted to relatively narrow niche modeling, paleoclimate models, and the early Cenozoic fossil geographic regions today were formerly found in different parts of record to examine the influence of climatic change on avian geo- – graphic distributions over the last ∼56 million years. -

Western North Greenland (Laurentia)
BULLETIN OF THE GEOLOGICAL SOCIETY OF DENMARK · VOL. 69 · 2021 Trilobite fauna of the Telt Bugt Formation (Cambrian Series 2–Miaolingian Series), western North Greenland (Laurentia) JOHN S. PEEL Peel, J.S. 2021. Trilobite fauna of the Telt Bugt Formation (Cambrian Series 2–Mi- aolingian Series), western North Greenland (Laurentia). Bulletin of the Geological Society of Denmark, Vol. 69, pp. 1–33. ISSN 2245-7070. https://doi.org/10.37570/bgsd-2021-69-01 Trilobites dominantly of middle Cambrian (Miaolingian Series, Wuliuan Stage) Geological Society of Denmark age are described from the Telt Bugt Formation of Daugaard-Jensen Land, western https://2dgf.dk North Greenland (Laurentia), which is a correlative of the Cape Wood Formation of Inglefield Land and Ellesmere Island, Nunavut. Four biozones are recognised in Received 6 July 2020 Daugaard-Jensen Land, representing the Delamaran and Topazan regional stages Accepted in revised form of the western USA. The basal Plagiura–Poliella Biozone, with Mexicella cf. robusta, 16 December 2020 Kochiella, Fieldaspis? and Plagiura?, straddles the Cambrian Series 2–Miaolingian Series Published online 20 January 2021 boundary. It is overlain by the Mexicella mexicana Biozone, recognised for the first time in Greenland, with rare specimens of Caborcella arrojosensis. The Glossopleura walcotti © 2021 the authors. Re-use of material is Biozone, with Glossopleura, Clavaspidella and Polypleuraspis, dominates the succes- permitted, provided this work is cited. sion in eastern Daugaard-Jensen Land but is seemingly not represented in the type Creative Commons License CC BY: section in western outcrops, likely reflecting the drastic thinning of the formation https://creativecommons.org/licenses/by/4.0/ towards the north-west. -

Guadalupian, Middle Permian) Mass Extinction in NW Pangea (Borup Fiord, Arctic Canada): a Global Crisis Driven by Volcanism and Anoxia
The Capitanian (Guadalupian, Middle Permian) mass extinction in NW Pangea (Borup Fiord, Arctic Canada): A global crisis driven by volcanism and anoxia David P.G. Bond1†, Paul B. Wignall2, and Stephen E. Grasby3,4 1Department of Geography, Geology and Environment, University of Hull, Hull, HU6 7RX, UK 2School of Earth and Environment, University of Leeds, Leeds, LS2 9JT, UK 3Geological Survey of Canada, 3303 33rd Street N.W., Calgary, Alberta, T2L 2A7, Canada 4Department of Geoscience, University of Calgary, 2500 University Drive N.W., Calgary Alberta, T2N 1N4, Canada ABSTRACT ing gun of eruptions in the distant Emeishan 2009; Wignall et al., 2009a, 2009b; Bond et al., large igneous province, which drove high- 2010a, 2010b), making this a mid-Capitanian Until recently, the biotic crisis that oc- latitude anoxia via global warming. Although crisis of short duration, fulfilling the second cri- curred within the Capitanian Stage (Middle the global Capitanian extinction might have terion. Several other marine groups were badly Permian, ca. 262 Ma) was known only from had different regional mechanisms, like the affected in equatorial eastern Tethys Ocean, in- equatorial (Tethyan) latitudes, and its global more famous extinction at the end of the cluding corals, bryozoans, and giant alatocon- extent was poorly resolved. The discovery of Permian, each had its roots in large igneous chid bivalves (e.g., Wang and Sugiyama, 2000; a Boreal Capitanian crisis in Spitsbergen, province volcanism. Weidlich, 2002; Bond et al., 2010a; Chen et al., with losses of similar magnitude to those in 2018). In contrast, pelagic elements of the fauna low latitudes, indicated that the event was INTRODUCTION (ammonoids and conodonts) suffered a later, geographically widespread, but further non- ecologically distinct, extinction crisis in the ear- Tethyan records are needed to confirm this as The Capitanian (Guadalupian Series, Middle liest Lopingian (Huang et al., 2019). -
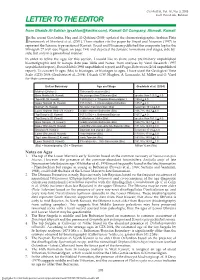
Letter to the Editor
GeoArabia, Vol. 10, No. 3, 2005 Gulf PetroLink, Bahrain LETTER TO THE EDITOR from Ghaida Al-Sahlan ([email protected]), Kuwait Oil Company, Ahmadi, Kuwait n the recent GeoArabia, Haq and Al-Qahtani (2005) updated the chronostratigraphic Arabian Plate Iframework of Sharland et al. (2001). These studies cite the paper by Yousif and Nouman (1997) to represent the Jurassic type section of Kuwait. Yousif and Nouman published the composite log for the Minagish-27 well (see Figure on page 194) and depicted the Jurassic formations and stages, side-by- side, but only in a generalized manner. In order to refine the ages for this section, I would like to share some preliminary unpublished biostratigraphic and Sr isotope data (see Table and Notes) from analyses by Varol Research (1997 unpublished report), ExxonMobil (1998 unpublished report) and Fugro-Robertson (2004 unpublished report). To convert Sr ages (Ma) to biostages, or biostages to ages, I have used the Geological Time Scale (GTS) 2004 (Gradstein et al., 2004). I thank G.W. Hughes, A. Lomando, M. Miller and O. Varol for their comments. Unit or Boundary Age and Stage Gradstein et al. (2004) Makhul (Offshore) Tithonian-Berriasian (Bio) Base Makhul (N. Kuwait) No younger than Tithonian (Bio) greater than 145.5 + 4.0 Top Hith (W. Kuwait) 150.0 (Sr) = c. Tithonian/Kimmeridgian ? 150.8 + 4.0 Upper Najmah (S. Kuwait) 155.0 (Sr) = c. Kimmeridgian/Oxfordian 155.7 + 4.0 Najmah (N. Kuwait) No older than Oxfordian (Bio) less than 161.2 + 4.0 Lower Najmah Shale (N. Kuwait) middle and late Bathonian (Bio) 166.7 to 164.7 + 4.0 Top Sargelu (S. -

A Comparison of the Dinosaur Communities from the Middle
Preprints (www.preprints.org) | NOT PEER-REVIEWED | Posted: 31 July 2018 doi:10.20944/preprints201807.0610.v1 Peer-reviewed version available at Geosciences 2018, 8, 327; doi:10.3390/geosciences8090327 1 Review 2 A comparison of the dinosaur communities from 3 the Middle Jurassic of the Cleveland (Yorkshire) 4 and Hebrides (Skye) basins, based on their ichnites 5 6 Mike Romano 1*, Neil D. L. Clark 2 and Stephen L. Brusatte 3 7 1 Independent Researcher, 14 Green Lane, Dronfield, Sheffield S18 2LZ, England, United Kingdom; 8 [email protected] 9 2 Curator of Palaeontology, The Hunterian, University of Glasgow, University Avenue, Glasgow 10 G12 8QQ, Scotland, United Kingdom; [email protected] 11 3 Chancellor's Fellow in Vertebrate Palaeontology, School of Geosciences, University of Edinburgh, 12 Grant Institute, The King's Buildings, James Hutton Road, Edinburgh EH9 3FE, Scotland, United Kingdom; 13 [email protected] 14 * Correspondence: [email protected]; Tel.: 01246 417330 15 16 Abstract: 17 Despite the Hebrides and Cleveland basins being geographically close, research has not 18 previously been carried out to determine faunal similarities and assess the possibility of links 19 between the dinosaur populations. The palaeogeography of both areas during the Middle Jurassic 20 shows that there were no elevated landmasses being eroded to produce conglomeratic material in 21 the basins at that time. The low-lying landscape and connected shorelines may have provided 22 connectivity between the two dinosaur populations. 23 The dinosaur fauna of the Hebrides and Cleveland basins has been assessed based primarily 24 on the abundant ichnites found in both areas as well as their skeletal remains. -

Evaluating the Frasnian-Famennian Mass Extinction: Comparing Brachiopod Faunas
Evaluating the Frasnian-Famennian mass extinction: Comparing brachiopod faunas PAUL COPPER Copper, P. 1998. Evaluating the Frasnian-Famennian mass extinction: Comparing bra- chiopod faunas.- Acta Palaeontologica Polonica 43,2,137-154. The Frasnian-Famennian (F-F) mass extinctions saw the global loss of all genera belonging to the tropically confined order Atrypida (and Pentamerida): though Famen- nian forms have been reported in the literafure, none can be confirmed. Losses were more severe during the Givetian (including the extinction of the suborder Davidsoniidina, and the reduction of the suborder Lissatrypidina to a single genus),but ońgination rates in the remaining suborder surviving into the Frasnian kept the group alive, though much reduced in biodiversity from the late Earb and Middle Devonian. In the terminal phases of the late Palmatolepis rhenana and P linguifurmis zones at the end of the Frasnian, during which the last few Atrypidae dechned, no new genera originated, and thus the Atrypida were extĘated. There is no evidence for an abrupt termination of all lineages at the F-F boundary, nor that the Atrypida were abundant at this time, since all groups were in decline and impoverished. Atypida were well established in dysaerobic, muddy substrate, reef lagoonal and off-reef deeper water settings in the late Givetian and Frasnian, alongside a range of brachiopod orders which sailed through the F-F boundary: tropical shelf anoxia or hypońa seems implausible as a cause for aĘpid extinction. Glacial-interglacial climate cycles recorded in South Ameńca for the Late Devonian, and their synchronous global cooling effect in low latitudes, as well as loss of the reef habitat and shelf area reduction, remain as the most likely combined scenarios for the mass extinction events. -

Lee-Riding-2018.Pdf
Earth-Science Reviews 181 (2018) 98–121 Contents lists available at ScienceDirect Earth-Science Reviews journal homepage: www.elsevier.com/locate/earscirev Marine oxygenation, lithistid sponges, and the early history of Paleozoic T skeletal reefs ⁎ Jeong-Hyun Leea, , Robert Ridingb a Department of Geology and Earth Environmental Sciences, Chungnam National University, Daejeon 34134, Republic of Korea b Department of Earth and Planetary Sciences, University of Tennessee, Knoxville, TN 37996, USA ARTICLE INFO ABSTRACT Keywords: Microbial carbonates were major components of early Paleozoic reefs until coral-stromatoporoid-bryozoan reefs Cambrian appeared in the mid-Ordovician. Microbial reefs were augmented by archaeocyath sponges for ~15 Myr in the Reef gap early Cambrian, by lithistid sponges for the remaining ~25 Myr of the Cambrian, and then by lithistid, calathiid Dysoxia and pulchrilaminid sponges for the first ~25 Myr of the Ordovician. The factors responsible for mid–late Hypoxia Cambrian microbial-lithistid sponge reef dominance remain unclear. Although oxygen increase appears to have Lithistid sponge-microbial reef significantly contributed to the early Cambrian ‘Explosion’ of marine animal life, it was followed by a prolonged period dominated by ‘greenhouse’ conditions, as sea-level rose and CO2 increased. The mid–late Cambrian was unusually warm, and these elevated temperatures can be expected to have lowered oxygen solubility, and to have promoted widespread thermal stratification resulting in marine dysoxia and hypoxia. Greenhouse condi- tions would also have stimulated carbonate platform development, locally further limiting shallow-water cir- culation. Low marine oxygenation has been linked to episodic extinctions of phytoplankton, trilobites and other metazoans during the mid–late Cambrian.