Abnormal Protein Phosphorylation in Human Amyotrophic Lateral Sclerosis
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Related Malignant Phenotypes in the Nf1-Deficient MPNST
Published OnlineFirst February 19, 2013; DOI: 10.1158/1541-7786.MCR-12-0593 Molecular Cancer Genomics Research RAS/MEK–Independent Gene Expression Reveals BMP2- Related Malignant Phenotypes in the Nf1-Deficient MPNST Daochun Sun1, Ramsi Haddad2,3, Janice M. Kraniak2, Steven D. Horne1, and Michael A. Tainsky1,2 Abstract Malignant peripheral nerve sheath tumor (MPNST) is a type of soft tissue sarcoma that occurs in carriers of germline mutations in Nf1 gene as well as sporadically. Neurofibromin, encoded by the Nf1 gene, functions as a GTPase-activating protein (GAP) whose mutation leads to activation of wt-RAS and mitogen-activated protein kinase (MAPK) signaling in neurofibromatosis type I (NF1) patients' tumors. However, therapeutic targeting of RAS and MAPK have had limited success in this disease. In this study, we modulated NRAS, mitogen-activated protein/extracellular signal–regulated kinase (MEK)1/2, and neurofibromin levels in MPNST cells and determined gene expression changes to evaluate the regulation of signaling pathways in MPNST cells. Gene expression changes due to neurofibromin modulation but independent of NRAS and MEK1/2 regulation in MPNST cells indicated bone morphogenetic protein 2 (Bmp2) signaling as a key pathway. The BMP2-SMAD1/5/8 pathway was activated in NF1-associated MPNST cells and inhibition of BMP2 signaling by LDN-193189 or short hairpin RNA (shRNA) to BMP2 decreased the motility and invasion of NF1-associated MPNST cells. The pathway-specific gene changes provide a greater understanding of the complex role of neurofibromin in MPNST pathology and novel targets for drug discovery. Mol Cancer Res; 11(6); 616–27. -

NICU Gene List Generator.Xlsx
Neonatal Crisis Sequencing Panel Gene List Genes: A2ML1 - B3GLCT A2ML1 ADAMTS9 ALG1 ARHGEF15 AAAS ADAMTSL2 ALG11 ARHGEF9 AARS1 ADAR ALG12 ARID1A AARS2 ADARB1 ALG13 ARID1B ABAT ADCY6 ALG14 ARID2 ABCA12 ADD3 ALG2 ARL13B ABCA3 ADGRG1 ALG3 ARL6 ABCA4 ADGRV1 ALG6 ARMC9 ABCB11 ADK ALG8 ARPC1B ABCB4 ADNP ALG9 ARSA ABCC6 ADPRS ALK ARSL ABCC8 ADSL ALMS1 ARX ABCC9 AEBP1 ALOX12B ASAH1 ABCD1 AFF3 ALOXE3 ASCC1 ABCD3 AFF4 ALPK3 ASH1L ABCD4 AFG3L2 ALPL ASL ABHD5 AGA ALS2 ASNS ACAD8 AGK ALX3 ASPA ACAD9 AGL ALX4 ASPM ACADM AGPS AMELX ASS1 ACADS AGRN AMER1 ASXL1 ACADSB AGT AMH ASXL3 ACADVL AGTPBP1 AMHR2 ATAD1 ACAN AGTR1 AMN ATL1 ACAT1 AGXT AMPD2 ATM ACE AHCY AMT ATP1A1 ACO2 AHDC1 ANK1 ATP1A2 ACOX1 AHI1 ANK2 ATP1A3 ACP5 AIFM1 ANKH ATP2A1 ACSF3 AIMP1 ANKLE2 ATP5F1A ACTA1 AIMP2 ANKRD11 ATP5F1D ACTA2 AIRE ANKRD26 ATP5F1E ACTB AKAP9 ANTXR2 ATP6V0A2 ACTC1 AKR1D1 AP1S2 ATP6V1B1 ACTG1 AKT2 AP2S1 ATP7A ACTG2 AKT3 AP3B1 ATP8A2 ACTL6B ALAS2 AP3B2 ATP8B1 ACTN1 ALB AP4B1 ATPAF2 ACTN2 ALDH18A1 AP4M1 ATR ACTN4 ALDH1A3 AP4S1 ATRX ACVR1 ALDH3A2 APC AUH ACVRL1 ALDH4A1 APTX AVPR2 ACY1 ALDH5A1 AR B3GALNT2 ADA ALDH6A1 ARFGEF2 B3GALT6 ADAMTS13 ALDH7A1 ARG1 B3GAT3 ADAMTS2 ALDOB ARHGAP31 B3GLCT Updated: 03/15/2021; v.3.6 1 Neonatal Crisis Sequencing Panel Gene List Genes: B4GALT1 - COL11A2 B4GALT1 C1QBP CD3G CHKB B4GALT7 C3 CD40LG CHMP1A B4GAT1 CA2 CD59 CHRNA1 B9D1 CA5A CD70 CHRNB1 B9D2 CACNA1A CD96 CHRND BAAT CACNA1C CDAN1 CHRNE BBIP1 CACNA1D CDC42 CHRNG BBS1 CACNA1E CDH1 CHST14 BBS10 CACNA1F CDH2 CHST3 BBS12 CACNA1G CDK10 CHUK BBS2 CACNA2D2 CDK13 CILK1 BBS4 CACNB2 CDK5RAP2 -

The Role of the Rho Gtpases in Neuronal Development
Downloaded from genesdev.cshlp.org on September 24, 2021 - Published by Cold Spring Harbor Laboratory Press REVIEW The role of the Rho GTPases in neuronal development Eve-Ellen Govek,1,2, Sarah E. Newey,1 and Linda Van Aelst1,2,3 1Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, 11724, USA; 2Molecular and Cellular Biology Program, State University of New York at Stony Brook, Stony Brook, New York, 11794, USA Our brain serves as a center for cognitive function and and an inactive GDP-bound state. Their activity is de- neurons within the brain relay and store information termined by the ratio of GTP to GDP in the cell and can about our surroundings and experiences. Modulation of be influenced by a number of different regulatory mol- this complex neuronal circuitry allows us to process that ecules. Guanine nucleotide exchange factors (GEFs) ac- information and respond appropriately. Proper develop- tivate GTPases by enhancing the exchange of bound ment of neurons is therefore vital to the mental health of GDP for GTP (Schmidt and Hall 2002); GTPase activat- an individual, and perturbations in their signaling or ing proteins (GAPs) act as negative regulators of GTPases morphology are likely to result in cognitive impairment. by enhancing the intrinsic rate of GTP hydrolysis of a The development of a neuron requires a series of steps GTPase (Bernards 2003; Bernards and Settleman 2004); that begins with migration from its birth place and ini- and guanine nucleotide dissociation inhibitors (GDIs) tiation of process outgrowth, and ultimately leads to dif- prevent exchange of GDP for GTP and also inhibit the ferentiation and the formation of connections that allow intrinsic GTPase activity of GTP-bound GTPases (Zalc- it to communicate with appropriate targets. -

Structural, Biochemical, and Cell Biological Characterization of Rab7 Mutants That Cause Peripheral Neuropathy
University of Pennsylvania ScholarlyCommons Publicly Accessible Penn Dissertations Spring 2010 Structural, Biochemical, and Cell Biological Characterization of Rab7 Mutants That Cause Peripheral Neuropathy Brett A. McCray University of Pennsylvania, [email protected] Follow this and additional works at: https://repository.upenn.edu/edissertations Part of the Molecular and Cellular Neuroscience Commons Recommended Citation McCray, Brett A., "Structural, Biochemical, and Cell Biological Characterization of Rab7 Mutants That Cause Peripheral Neuropathy" (2010). Publicly Accessible Penn Dissertations. 145. https://repository.upenn.edu/edissertations/145 This paper is posted at ScholarlyCommons. https://repository.upenn.edu/edissertations/145 For more information, please contact [email protected]. Structural, Biochemical, and Cell Biological Characterization of Rab7 Mutants That Cause Peripheral Neuropathy Abstract Coordinated trafficking of intracellular vesicles is of critical importance for the maintenance of cellular health and homeostasis. Members of the Rab GTPase family serve as master regulators of vesicular trafficking, maturation, and fusion by reversibly associating with distinct target membranes and recruiting specific effector proteins. Rabs act as molecular switches by cycling between an active, GTP-bound form and an inactive, GDP-bound form. The activity cycle is coupled to GTP hydrolysis and is tightly controlled by regulatory proteins such as guanine nucleotide exchange factors and GTPase activating proteins. Rab7 specifically regulates the trafficking and maturation of vesicle populations that are involved in protein degradation including late endosomes, lysosomes, and autophagic vacuoles. Missense mutations of Rab7 cause a dominantly-inherited axonal degeneration known as Charcot-Marie-Tooth type 2B (CMT2B) through an unknown mechanism. Patients with CMT2B present with length-dependent degeneration of peripheral sensory and motor neurons that leads to weakness and profound sensory loss. -

Appendix 2. Significantly Differentially Regulated Genes in Term Compared with Second Trimester Amniotic Fluid Supernatant
Appendix 2. Significantly Differentially Regulated Genes in Term Compared With Second Trimester Amniotic Fluid Supernatant Fold Change in term vs second trimester Amniotic Affymetrix Duplicate Fluid Probe ID probes Symbol Entrez Gene Name 1019.9 217059_at D MUC7 mucin 7, secreted 424.5 211735_x_at D SFTPC surfactant protein C 416.2 206835_at STATH statherin 363.4 214387_x_at D SFTPC surfactant protein C 295.5 205982_x_at D SFTPC surfactant protein C 288.7 1553454_at RPTN repetin solute carrier family 34 (sodium 251.3 204124_at SLC34A2 phosphate), member 2 238.9 206786_at HTN3 histatin 3 161.5 220191_at GKN1 gastrokine 1 152.7 223678_s_at D SFTPA2 surfactant protein A2 130.9 207430_s_at D MSMB microseminoprotein, beta- 99.0 214199_at SFTPD surfactant protein D major histocompatibility complex, class II, 96.5 210982_s_at D HLA-DRA DR alpha 96.5 221133_s_at D CLDN18 claudin 18 94.4 238222_at GKN2 gastrokine 2 93.7 1557961_s_at D LOC100127983 uncharacterized LOC100127983 93.1 229584_at LRRK2 leucine-rich repeat kinase 2 HOXD cluster antisense RNA 1 (non- 88.6 242042_s_at D HOXD-AS1 protein coding) 86.0 205569_at LAMP3 lysosomal-associated membrane protein 3 85.4 232698_at BPIFB2 BPI fold containing family B, member 2 84.4 205979_at SCGB2A1 secretoglobin, family 2A, member 1 84.3 230469_at RTKN2 rhotekin 2 82.2 204130_at HSD11B2 hydroxysteroid (11-beta) dehydrogenase 2 81.9 222242_s_at KLK5 kallikrein-related peptidase 5 77.0 237281_at AKAP14 A kinase (PRKA) anchor protein 14 76.7 1553602_at MUCL1 mucin-like 1 76.3 216359_at D MUC7 mucin 7, -

Immersion Course Lecture Kelly
Genomic Counseling in the research setting Kelly East, MS CGC Genetic Counselor HudsonAlpha Institute for Biotechnology single gene disorders • mutation in a single gene leads to disease • often has characteristic family inheritance patterns J Neurol Neurosurg Psychiatry 2002;73:ii5-ii11 Genes that code for hemoglobin 1 2 3 4 5 HBB 6 7 8 9 10 11 12 HBA2 13 14 15 16 17 18 19 20 21 22 X Y http://www.genome.gov/glossary/resources/karyotype.pdf TARDBP Genes associated ALS2 with ALS 1 2 3 4 5 C9ORF72 OPTN FIG4 ATXN2 SETX 6 7 8 9 10 11 12 ANG SPG11 FUS 13 14 15 16 17 18 VAPB UBQLN2 SOD1 19 20 21 22 X Y http://www.genome.gov/glossary/resources/karyotype.pdf complex traits • most common traits and diseases have a complex etiology • causative risks can include genetic changes (both large and small scale) environmental factors (head injury, nutrition, exposure to toxins) societal factors (death of family member, abuse, hardships) • in most cases - not triggered by a change in a single gene but rather by the interaction of several genetic, environmental and societal risks spectrum of human genetic conditions CGTATACCGGGTCATGCACGTGTAGAGCGAGTTAGCTCGCTGGCTAAAGAGGGTCGAC ATCCGCGAGTTTATGAGGAAGAATCGGCAGCTTGACCGAAGAGGCGTGGTAAGACCCG TTAGGGATCGTATACCGGGTCATGCACGTGTAGAGCGAGTTAGCTCGCTGGCTAAAGA GGGTCGACATCCGCGAGTTTATGAGGAAGAATCGGCAGCTTGACCGAAGAGGCGTGGT AAGACCCGTTAGGGATCGTATACCGGGTCATGCACGTGTAGAGCGAGTTAGCTCGCTG GCTAAAGAGGGTCGACATCCGCGAGTTTATGAGGAAGAATCGGCAGCTTGACCGAAGA GGCGTGGTAAGACCCGTTAGGGATCGTATACCGGGTCATGCACGTGTAGAGCGAGTTA in patients with a suspected genetic -

Figure S1. HAEC ROS Production and ML090 NOX5-Inhibition
Figure S1. HAEC ROS production and ML090 NOX5-inhibition. (a) Extracellular H2O2 production in HAEC treated with ML090 at different concentrations and 24 h after being infected with GFP and NOX5-β adenoviruses (MOI 100). **p< 0.01, and ****p< 0.0001 vs control NOX5-β-infected cells (ML090, 0 nM). Results expressed as mean ± SEM. Fold increase vs GFP-infected cells with 0 nM of ML090. n= 6. (b) NOX5-β overexpression and DHE oxidation in HAEC. Representative images from three experiments are shown. Intracellular superoxide anion production of HAEC 24 h after infection with GFP and NOX5-β adenoviruses at different MOIs treated or not with ML090 (10 nM). MOI: Multiplicity of infection. Figure S2. Ontology analysis of HAEC infected with NOX5-β. Ontology analysis shows that the response to unfolded protein is the most relevant. Figure S3. UPR mRNA expression in heart of infarcted transgenic mice. n= 12-13. Results expressed as mean ± SEM. Table S1: Altered gene expression due to NOX5-β expression at 12 h (bold, highlighted in yellow). N12hvsG12h N18hvsG18h N24hvsG24h GeneName GeneDescription TranscriptID logFC p-value logFC p-value logFC p-value family with sequence similarity NM_052966 1.45 1.20E-17 2.44 3.27E-19 2.96 6.24E-21 FAM129A 129. member A DnaJ (Hsp40) homolog. NM_001130182 2.19 9.83E-20 2.94 2.90E-19 3.01 1.68E-19 DNAJA4 subfamily A. member 4 phorbol-12-myristate-13-acetate- NM_021127 0.93 1.84E-12 2.41 1.32E-17 2.69 1.43E-18 PMAIP1 induced protein 1 E2F7 E2F transcription factor 7 NM_203394 0.71 8.35E-11 2.20 2.21E-17 2.48 1.84E-18 DnaJ (Hsp40) homolog. -

Identification of Transcriptional Mechanisms Downstream of Nf1 Gene Defeciency in Malignant Peripheral Nerve Sheath Tumors Daochun Sun Wayne State University
Wayne State University DigitalCommons@WayneState Wayne State University Dissertations 1-1-2012 Identification of transcriptional mechanisms downstream of nf1 gene defeciency in malignant peripheral nerve sheath tumors Daochun Sun Wayne State University, Follow this and additional works at: http://digitalcommons.wayne.edu/oa_dissertations Recommended Citation Sun, Daochun, "Identification of transcriptional mechanisms downstream of nf1 gene defeciency in malignant peripheral nerve sheath tumors" (2012). Wayne State University Dissertations. Paper 558. This Open Access Dissertation is brought to you for free and open access by DigitalCommons@WayneState. It has been accepted for inclusion in Wayne State University Dissertations by an authorized administrator of DigitalCommons@WayneState. IDENTIFICATION OF TRANSCRIPTIONAL MECHANISMS DOWNSTREAM OF NF1 GENE DEFECIENCY IN MALIGNANT PERIPHERAL NERVE SHEATH TUMORS by DAOCHUN SUN DISSERTATION Submitted to the Graduate School of Wayne State University, Detroit, Michigan in partial fulfillment of the requirements for the degree of DOCTOR OF PHILOSOPHY 2012 MAJOR: MOLECULAR BIOLOGY AND GENETICS Approved by: _______________________________________ Advisor Date _______________________________________ _______________________________________ _______________________________________ © COPYRIGHT BY DAOCHUN SUN 2012 All Rights Reserved DEDICATION This work is dedicated to my parents and my wife Ze Zheng for their continuous support and understanding during the years of my education. I could not achieve my goal without them. ii ACKNOWLEDGMENTS I would like to express tremendous appreciation to my mentor, Dr. Michael Tainsky. His guidance and encouragement throughout this project made this dissertation come true. I would also like to thank my committee members, Dr. Raymond Mattingly and Dr. John Reiners Jr. for their sustained attention to this project during the monthly NF1 group meetings and committee meetings, Dr. -

Rho Guanine Nucleotide Exchange Factors: Regulators of Rho Gtpase Activity in Development and Disease
Oncogene (2014) 33, 4021–4035 & 2014 Macmillan Publishers Limited All rights reserved 0950-9232/14 www.nature.com/onc REVIEW Rho guanine nucleotide exchange factors: regulators of Rho GTPase activity in development and disease DR Cook1, KL Rossman2,3 and CJ Der1,2,3 The aberrant activity of Ras homologous (Rho) family small GTPases (20 human members) has been implicated in cancer and other human diseases. However, in contrast to the direct mutational activation of Ras found in cancer and developmental disorders, Rho GTPases are activated most commonly in disease by indirect mechanisms. One prevalent mechanism involves aberrant Rho activation via the deregulated expression and/or activity of Rho family guanine nucleotide exchange factors (RhoGEFs). RhoGEFs promote formation of the active GTP-bound state of Rho GTPases. The largest family of RhoGEFs is comprised of the Dbl family RhoGEFs with 70 human members. The multitude of RhoGEFs that activate a single Rho GTPase reflects the very specific role of each RhoGEF in controlling distinct signaling mechanisms involved in Rho activation. In this review, we summarize the role of Dbl RhoGEFs in development and disease, with a focus on Ect2 (epithelial cell transforming squence 2), Tiam1 (T-cell lymphoma invasion and metastasis 1), Vav and P-Rex1/2 (PtdIns(3,4,5)P3 (phosphatidylinositol (3,4,5)-triphosphate)-dependent Rac exchanger). Oncogene (2014) 33, 4021–4035; doi:10.1038/onc.2013.362; published online 16 September 2013 Keywords: Rac1; RhoA; Cdc42; guanine nucleotide exchange factors; cancer; -
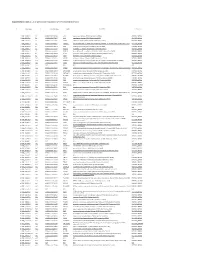
Supplemetary Table 2. List of Genes Down-Regulated in LPAR6 Knocked Down Cells
Supplemetary Table 2. List of genes down-regulated in LPAR6 knocked down cells g# initial alias c# converted alias name description namespace 1 NM_002317.5 1.1 ENSG00000113083 LOX lysyl oxidase [Source:HGNC Symbol;Acc:6664] REFSEQ_MRNA 2 NM_006183.4 2.1 ENSG00000133636 NTS neurotensin [Source:HGNC Symbol;Acc:8038] REFSEQ_MRNA 3 NM_005213.3 3.1 ENSG00000121552 CSTA cystatin A (stefin A) [Source:HGNC Symbol;Acc:2481] REFSEQ_MRNA 4 NM_007231.3 4.1 ENSG00000087916 SLC6A14 solute carrier family 6 (amino acid transporter), member 14 [Source:HGNC Symbol;Acc:11047] REFSEQ_MRNA 5 NM_001873.2 5.1 ENSG00000109472 CPE carboxypeptidase E [Source:HGNC Symbol;Acc:2303] REFSEQ_MRNA 6 NM_019856.1 6.1 ENSG00000101605 MYOM1 myomesin 1, 185kDa [Source:HGNC Symbol;Acc:7613] REFSEQ_MRNA 7 NM_032590.4 7.1 ENSG00000089094 KDM2B lysine (K)-specific demethylase 2B [Source:HGNC Symbol;Acc:13610] REFSEQ_MRNA 8 NM_001901.2 8.1 ENSG00000118523 CTGF connective tissue growth factor [Source:HGNC Symbol;Acc:2500] REFSEQ_MRNA 9 NM_007183.2 9.1 ENSG00000184363 PKP3 plakophilin 3 [Source:HGNC Symbol;Acc:9025] REFSEQ_MRNA 10 NM_182965.2 10.1 ENSG00000176170 SPHK1 sphingosine kinase 1 [Source:HGNC Symbol;Acc:11240] REFSEQ_MRNA 11 NM_152423.4 11.1 ENSG00000157502 MUM1L1 melanoma associated antigen (mutated) 1-like 1 [Source:HGNC Symbol;Acc:26583] REFSEQ_MRNA 12 NM_002923.3 12.1 ENSG00000116741 RGS2 regulator of G-protein signaling 2, 24kDa [Source:HGNC Symbol;Acc:9998] REFSEQ_MRNA 13 NR_003038.2 13.1 N/A N/A N/A N/A 14 NM_080862.1 14.1 ENSG00000175093 SPSB4 splA/ryanodine receptor -

Linkage of the Gene for an Autosomal Dominant Form of Juvenile Amyotrophic Lateral Sclerosis to Chromosome 9Q34 Phillip F
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Elsevier - Publisher Connector Am. J. Hum. Genet. 62:633–640, 1998 Linkage of the Gene for an Autosomal Dominant Form of Juvenile Amyotrophic Lateral Sclerosis to Chromosome 9q34 Phillip F. Chance,1 Bruce A. Rabin,2 Stephen G. Ryan,1 Yuan Ding,1 Mena Scavina,1 Barbara Crain,3 John W. Griffin,2,4 and David R. Cornblath2 1Division of Neurology, Children’s Hospital of Philadelphia, and Departments of Neurology and Pediatrics, University of Pennsylvania School of Medicine, Philadelphia; and Departments of 2Neurology, 3Pathology, and 4Neuroscience, Johns Hopkins University School of Medicine, Baltimore Summary of patients have similarly affected relatives; this indicates that genetic factors play an important role in the de- We performed genetic mapping studies of an 11-gen- velopment of some motor-neuron disorders (Strong et eration pedigree with an autosomal dominant, juvenile- al. 1991). Inherited forms of ALS are classified as “fa- onset motor-systems disease. The disorder is character- milial ALS” (Mulder et al. 1986). The clinical pheno- ized by slow progression, distal limb amyotrophy, and types of sporadic and familial ALS can be virtually in- pyramidal tract signs associated with severe loss of mo- distinguishable; however, pathological studies in tor neurons in the brain stem and spinal cord. The gene pedigrees with familial ALS have documented more ex- for this disorder, classified as a form of juvenile amy- tensive, subclinical involvement of the nervous system, otrophic lateral sclerosis (ALS), is designated “ALS4.” including degeneration of the posterior columns, We performed a genomewide search and detected strong Clarke’s column, and spinocerebellar tracts (Engel et al. -

Investigation of Two Early Events in Amyotrophic Lateral Sclerosis
INVESTIGATION OF TWO EARLY EVENTS IN AMYOTROPHIC LATERAL SCLEROSIS -MRNA OXIDATION AND UP-REGULATION OF A NOVEL PROTECTIVE FACTOR MSUR1 DISSERTATION Presented in Partial Fulfillment of the Requirements for the Degree Doctor of Philosophy in the Graduate School of The Ohio State University By Yueming Chang, Bachelor in Medicine *********** The Ohio State University 2007 Dissertation Committee: Approved by Professor Chien-liang Glenn Lin, Advisor Professor Andrej Rotter ------------------------------------------- Professor Arthur H.M. Burghes Advisor Professor John Oberdick The Ohio State Biochemistry Program ABSTRACT Amyotrophic Lateral Sclerosis (ALS) is a fatal neurodegenerative disorder that is characterized by progressive degeneration of motor neurons in the spinal cord, motor cortex and brainstem, which typically results in mortality within 2-5 years after the onset of disease. The cause of disease is unknown in the majority of cases, and there is no cure for ALS of the moment. Approximately 5% of ALS cases are familial, and 15-25% of the familial cases are linked to mutation in the gene encoding the antioxidant enzyme Cu 2+ /Zn 2+ superoxide dismutase (SOD1). Overexpression of some of ALS-linked mutant SOD1 proteins in transgenic mice results in the development of a neurological disorder that resembles ALS patients. The transgenic mice expressing mutant SOD1 (G93A) is a commonly used ALS animal model. This dissertation demonstrates two early events occurred in the pre-symptomatic stage of SOD1 (G93A) mice, including RNA oxidation and up-regulation of a novel protective factor MSUR1 (mutant SOD1-upregulated RNA 1). Accumulating evidence indicates that messenger RNA (mRNA) oxidation may be associated with neuronal deterioration during the process of neurodegeneration.