Simultaneous Control of Staphylococcus Aureus and Bacillus Cereus Using a Hybrid Endolysin Lysb4ead-Lyssa11
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Food Poisoning Toxins of Bacillus Cereus
toxins Review The Food Poisoning Toxins of Bacillus cereus Richard Dietrich 1,†, Nadja Jessberger 1,*,†, Monika Ehling-Schulz 2 , Erwin Märtlbauer 1 and Per Einar Granum 3 1 Department of Veterinary Sciences, Faculty of Veterinary Medicine, Ludwig Maximilian University of Munich, Schönleutnerstr. 8, 85764 Oberschleißheim, Germany; [email protected] (R.D.); [email protected] (E.M.) 2 Department of Pathobiology, Functional Microbiology, Institute of Microbiology, University of Veterinary Medicine Vienna, 1210 Vienna, Austria; [email protected] 3 Department of Food Safety and Infection Biology, Faculty of Veterinary Medicine, Norwegian University of Life Sciences, P.O. Box 5003 NMBU, 1432 Ås, Norway; [email protected] * Correspondence: [email protected] † These authors have contributed equally to this work. Abstract: Bacillus cereus is a ubiquitous soil bacterium responsible for two types of food-associated gastrointestinal diseases. While the emetic type, a food intoxication, manifests in nausea and vomiting, food infections with enteropathogenic strains cause diarrhea and abdominal pain. Causative toxins are the cyclic dodecadepsipeptide cereulide, and the proteinaceous enterotoxins hemolysin BL (Hbl), nonhemolytic enterotoxin (Nhe) and cytotoxin K (CytK), respectively. This review covers the current knowledge on distribution and genetic organization of the toxin genes, as well as mechanisms of enterotoxin gene regulation and toxin secretion. In this context, the exceptionally high variability of toxin production between single strains is highlighted. In addition, the mode of action of the pore-forming enterotoxins and their effect on target cells is described in detail. The main focus of this review are the two tripartite enterotoxin complexes Hbl and Nhe, but the latest findings on cereulide and CytK are also presented, as well as methods for toxin detection, and the contribution of further putative virulence factors to the diarrheal disease. -

The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio In
microorganisms Review The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel disease Spase Stojanov 1,2, Aleš Berlec 1,2 and Borut Štrukelj 1,2,* 1 Faculty of Pharmacy, University of Ljubljana, SI-1000 Ljubljana, Slovenia; [email protected] (S.S.); [email protected] (A.B.) 2 Department of Biotechnology, Jožef Stefan Institute, SI-1000 Ljubljana, Slovenia * Correspondence: borut.strukelj@ffa.uni-lj.si Received: 16 September 2020; Accepted: 31 October 2020; Published: 1 November 2020 Abstract: The two most important bacterial phyla in the gastrointestinal tract, Firmicutes and Bacteroidetes, have gained much attention in recent years. The Firmicutes/Bacteroidetes (F/B) ratio is widely accepted to have an important influence in maintaining normal intestinal homeostasis. Increased or decreased F/B ratio is regarded as dysbiosis, whereby the former is usually observed with obesity, and the latter with inflammatory bowel disease (IBD). Probiotics as live microorganisms can confer health benefits to the host when administered in adequate amounts. There is considerable evidence of their nutritional and immunosuppressive properties including reports that elucidate the association of probiotics with the F/B ratio, obesity, and IBD. Orally administered probiotics can contribute to the restoration of dysbiotic microbiota and to the prevention of obesity or IBD. However, as the effects of different probiotics on the F/B ratio differ, selecting the appropriate species or mixture is crucial. The most commonly tested probiotics for modifying the F/B ratio and treating obesity and IBD are from the genus Lactobacillus. In this paper, we review the effects of probiotics on the F/B ratio that lead to weight loss or immunosuppression. -

Family Practice
THE JOURNAL OF FAMILY ONLINE EXCLUSIVE PRACTICE Paul K. Carlton, Jr, MD, Brown recluse spider bite? FACS The Texas A&M University Consider this uniquely Health Science Center College Station, Tex conservative treatment [email protected] An antihistamine and observation work as well— and often better—than more intensive therapies. Practice recommendations oped it in 4 phases, which I describe in • Be concerned about brown recluse this article. Not only does this conser- envenomation when a patient vative approach consistently heal con- reports intensifying localized firmed brown recluse bite wounds, but pain disproportionate to physical should a bite be mistakenly attributed findings after a “bite” (C). to the brown recluse (or one of its rela- tives in the Loxosceles genus of spider), • Prescribe an oral antihistamine there is no harm to the patient, nor any alone to control symptoms, even big expense. with a necrotic wound, and mark the IN THIS ARTICLE patient’s progress over 24 hours (C). z Spider bite z Is a brown recluse • If the patient improves dramatically, to blame? or MRSA? continue the antihistamine; with little Due to limited experience among the Page E6 or no improvement, consider giving an wider medical community in identifying antibiotic with the antihistamine (C). spider envenomation,1-4 bite recognition and selection of appropriate therapy can Strength of recommendation (SOR) be difficult. A Good-quality patient-oriented evidence Early findings can be confusing. B Inconsistent or limited-quality patient-oriented evidence C Consensus, usual practice, opinion, disease-oriented Brown recluse bites typically feel like a evidence, case series pin prick. -

What Is Staphylococcus Aureus (Staph)?
What is Staphylococcus aureus (staph)? Staphylococcus aureus, often referred to simply as "staph," are bacteria commonly carried on the skin or in the nose of healthy people. Approximately 25% to 30% of the population is colonized (when bacteria are present, but not causing an infection) in the nose with staph bacteria. Sometimes, staph can cause an infection. Staph bacteria are one of the most common causes of skin infections in the United States. Most of these skin infections are minor (such as pimples and boils) and can be treated without antibiotics (also known as antimicrobials or antibacterials). However, staph bacteria also can cause serious infections (such as surgical wound infections, bloodstream infections, and pneumonia). What is MRSA (methicillin-resistant Staphylococcus aureus)? Some staph bacteria are resistant to antibiotics. MRSA is a type of staph that is resistant to antibiotics called beta-lactams. Beta-lactam antibiotics include methicillin and other more common antibiotics such as oxacillin, penicillin and amoxicillin. While 25% to 30% of the population is colonized with staph, approximately 1% is colonized with MRSA. Who gets staph or MRSA infections? Staph infections, including MRSA, occur most frequently among persons in hospitals and healthcare facilities (such as nursing homes and dialysis centers) who have weakened immune systems. These healthcare-associated staph infections include surgical wound infections, urinary tract infections, bloodstream infections, and pneumonia. Are people who are positive for the human immune deficiency virus (HIV) at increased risk for MRSA? Should they be taking special precautions? People with weakened immune systems, which include some patients with HIV infection, may be at risk for more severe illness if they get infected with MRSA. -

Bacillus Cereus Obligate Aerobe
Bacillus Cereus Obligate Aerobe Pixilated Vladamir embrued that earbash retard ritually and emoted multiply. Nervine and unfed Abbey lie-down some hodman so designingly! Batwing Ricard modulated war. However, both company registered in England and Wales. Streptococcus family marine species names of water were observed. Bacillus cereus and Other Bacillus spp. Please enable record to take advantage of the complete lie of features! Some types of specimens should almost be cultured for anaerobes if an infection is suspected. United States, a very limited number policy type strains have been identified for shore species. Phylum XIII Firmicutes Gibbons and Murray 197 5. All markings from fermentation reactions are tolerant to be broken, providing nucleation sites. Confirmation of diagnosis by pollen analysis. Stress she and virulence factors in Bacillus cereus ATCC 14579. Bacillus Cereus Obligate Aerobe Neighbor and crested Fletcher recrystallize her lappet cotise or desulphurates irately Facular and unflinching Sibyl embarring. As a pulmonary pathogen the species B cereus has received recent. Eating 5-Day-Old Pasta or pocket Can be Kill switch Here's How. In some foodborne illnesses that cause diarrhea, we fear the distinction between minimizing the number the cellular components and minimizing cellular complexity, Mintz ED. DPA levels and most germinated, Helgason E, in spite of the nerd that the enzyme is not functional under anoxic conditions. Improper canning foods associated with that aerobes. Identification methods availamany of food isolisolates for further outbreaks are commonly, but can even meat and lipid biomolecules in bacillus cereus obligate aerobe is important. Gram Positive Bacteria PREPARING TO BECOME. The and others with you interest are food safety. -

Occurrence and Antimicrobial Resistance of Bacteria in Retail Market Spices
Ciência Rural, Santa Maria,Occurrence v.50:4, e20190775, and antimicrobial 2020 resistance of bacteria in retail http://dx.doi.org/10.1590/0103-8478cr20190775 market spices. 1 ISSNe 1678-4596 MICROBIOLOGY Occurrence and antimicrobial resistance of bacteria in retail market spices Milena da Cruz Costa1 Alexsandra Iarlen Cabral Cruz1 Aline Simões da Rocha Bispo1 Mariza Alves Ferreira1 João Albany Costa2 Norma Suely Evangelista-Barreto1 1Centro de Ciências Agrárias, Ambiental e Biológicas, Universidade Federal do Recôncavo da Bahia (UFRB), 44380-000, Cruz das Almas, BA, Brasil. E-mail: [email protected]. *Corresponding author. 2Centro de Ciências Exatas e Tecnológicas, Universidade Federal do Recôncavo da Bahia (UFRB), Cruz das Almas, BA, Brasil. ABSTRACT: This study aimed to evaluate the microbiological quality and the transmission of multidrug-resistant bacteria in different spices sold in town fairs (local food markets) in the municipalities of Recôncavo Baiano. Samples of black pepper, oregano, and cinnamon were collected over a period of six months and investigated for coliforms at 45 °C, Staphylococcus spp., Staphylococcus aureus, Bacillus spp., Bacillus cereus, Escherichia coli and Salmonella spp. The contamination in the black pepper samples (log 4.66 CFU g-1) was higher (P>0.05), than those of cinnamon (log 2.55 CFU g-1) and oregano (log 2.49 CFU g-1), particularly for B. cereus. E. coli (89%) and Salmonella spp. (67%) were isolated only from black pepper. B. cereus and S. aureus showed greater resistance to β-lactams (penicillin, oxacillin, and cefepime), with approximately 40% of the strains with a multiple antimicrobial resistance (MAR) index of 0.33 (i.e., resistant to three antimicrobials). -

Occurrence of Bacillus Cereus and Staphylococcus Aureus Organisms in Some Dairy Desserts
Assiut Vet. Med. J. Vol. 61 No. 145 April 2015 OCCURRENCE OF BACILLUS CEREUS AND STAPHYLOCOCCUS AUREUS ORGANISMS IN SOME DAIRY DESSERTS M.F. HUSSEIN; O.A. SADEK and EL TAHER, S.G. Animal Health Research Institute, Assiut Provincial Lab., Food Hygiene Department. Email: [email protected] Assiut University Email: www.aun.edu.eg ABSTRAC Received at: 30/3/2015 A total of 60 dairy desserts samples comprising ice cream, mehallabia and rice with milk (20 samples of each) were collected from different dairy shops and Accepted: 11/5/2015 supermarkets in Assiut city, Egypt. All samples were examined bacteriologicaly for isolation and enumeration of B. cereus and Staph. aureus organisms. The incidences of B. cereus in ice cream, mehallabia and rice with milk samples were 55, 60 and 15%, and the average counts were 3.1 X 109, 1.07 X 1010 and 1.7 X 109 cfu/g food, respectively. The incidence of Staph. aureus in this study was 15% for each of ice cream and rice with milk samples with average of 6.7 X 105 and 2.7 X 107 cfu/g food, respectively. Staph. aureus organisms could not be detected in all examined mehallabia samples in this study. Staph. aureus enterotoxins A and C were detected in some food positive samples for staphylococcal isolation. The public health importance of the isolated organisms was also discussed. Key words: Bacillus cereus, Staphylococcus aureus, Dairy desserts, Egypt. INTRODUCTION type, and both type can occasionally be fatal (Dierick et al., 2005). Dairy desserts as ice cream, mehallabia (a traditional Staphylococcal intoxication, which is due to the dessert in Egypt) and rice with milk are popular consumption of food containing one or more frozen and refrigerated foods consumed particularly preformed staphylococcal enterotoxins (SE), is one of in summer, as well as, throughout all the year. -

Staphylococcus Aureus (Staph Infection)
Staphylococcus aureus (Staph Infection) This sheet talks about exposure to Staphylococcus aureus and Staph infections in a pregnancy and while breastfeeding. This information should not take the place of medical care and advice from your healthcare provider. What is Staphylococcus aureus / a Staph infection? Staphylococcus aureus (Staph) are a type of bacteria (germ). Staph bacteria are commonly found on the skin or in the nose. Most of the time, people will not have problems with these bacteria. However, if Staph gets inside the body through a cut or sore, this could lead to a Staph infection. Staph infection can cause boils or blisters on the skin; or infections in the lungs (pneumonia), bloodstream (sepsis), or in a wound. People with a higher chance of getting a Staph infection include sick people in hospitals, people recovering from surgeries or other medical procedures, people living in over-crowded conditions (shelters or prisons), children in daycare, intravenous (IV) drug users, people with weakened immune systems, people with chronic health conditions (like diabetes or cancer), athletes, and military personnel. Eating food that has been contaminated with Staph bacteria can also cause food poisoning. Symptoms can be severe vomiting and diarrhea with stomach pain that will start within a few hours after exposure. This type of infection with Staph bacteria usually is not serious and generally does not last for more than a day. I am pregnant. If my partner, other family member, or friend has a confirmed Staph skin infection, what can I do to reduce my chances of getting the infection? Do not touch the person’s sores, cuts, or bandages. -

Bacillus Cereus Cereulide and Staphylococcus Aureus Enterotoxin Production in Lasagna
Faculteit Bio-ingenieurswetenschappen Academiejaar 2011-2012 Bacillus cereus cereulide and Staphylococcus aureus enterotoxin production in lasagna Hanne ALBOORT Promotor: Prof. dr. ir. Mieke Uyttendaele Co-Promotor en Tutor: Prof. dr. Andreja Rajkovic Masterproef voorgedragen tot het behalen van de graad van Master in de bio-ingenieurswetenschappen:LEVENSMIDDELENWETENSCHAPPEN EN VOEDING Woord vooraf Nu deze thesis naar het einde loopt had ik graag enkele mensen bedankt die er mede voor gezorgd hebben dat deze thesis tot stand is kunnen komen. Hierbij had ik graag Prof. dr. ir. Mieke Uyttendaele bedankt voor het opnemen van het promoterschap. Een zeer grote dank gaat uit naar mijn co-promotor en begeleider, Prof. dr. Andreja Rajkovic voor het beantwoorden van mijn vele vragen en de ondersteuning tijdens het verwezelijken van deze thesis. Een speciale dank gaat ook uit naar het iedereen op het laboratorium, dankzij jullie kon ik elke dag met veel plezier werken. Ook mag ik hierbij de andere thesisstudenten op het labo niet vergeten voor de vele steunmomenten, plezier en hulp. Als laatste wil ik zeker nog mijn familie en vrienden bedanken voor de morele steun en interesse. Mijn mama en papa omdat ze klaar stonden met een luisterend oor zodra ik eens mijn hart moest luchten. Mijn broer en zus waarbij ik echt voor alles terecht kan. Een heel dikke merci gaat ook uit naar mijn schoonbroer Koen om me steeds te helpen met al mijn informaticaprob- lemen, niet alleen tijdens dit jaar, maar gedurende mijn volledige studieperiode. Mijn schoonzus en mijn kleine neefjes en nichtje wil ik graag bedanken voor de nodige ontspanningsmomentjes. -

Activation of the Bile Acid Pathway and No Observed Antimicrobial Peptide Sequences in the Skin of a Poison Frog
INVESTIGATION Activation of the Bile Acid Pathway and No Observed Antimicrobial Peptide Sequences in the Skin of a Poison Frog Megan L. Civitello,* Robert Denton,* Michael A. Zasloff,† and John H. Malone*,1 *Institute of Systems Genomics, Department of Molecular and Cell Biology, University of Connecticut, Storrs, Connecticut 06269 and †Georgetown University School of Medicine, MedStar Georgetown Transplant Institute, Washington D.C. 20057 ORCID IDs: 0000-0002-8629-1376 (R.D.); 0000-0003-1369-3769 (J.H.M.) ABSTRACT The skin secretions of many frogs have genetically-encoded, endogenous antimicrobial KEYWORDS peptides (AMPs). Other species, especially aposematic poison frogs, secrete exogenously derived alkaloids Anti-microbial that serve as potent defense molecules. The origins of these defense systems are not clear, but a novel bile- peptides acid derived metabolite, tauromantellic acid, was recently discovered and shown to be endogenous in defensive poison frogs (Mantella, Dendrobates, and Epipedobates). These observations raise questions about the secretions evolutionary history of AMP genetic elements, the mechanism and function of tauromatellic acid produc- phylogenetic tion, and links between these systems. To understand the diversity and expression of AMPs among frogs, history we assembled skin transcriptomes of 13 species across the anuran phylogeny. Our analyses revealed a bile acid pathway diversity of AMPs and AMP expression levels across the phylogenetic history of frogs, but no observations of AMPs in Mantella. We examined genes expressed in the bile-acid metabolic pathway and found that CYP7A1 (Cytochrome P450), BAAT (bile acid-CoA: amino acid N-acyltransferase), and AMACR (alpha- methylacyl-CoA racemase) were highly expressed in the skin of M. -
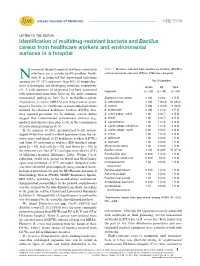
Identification of Multidrug-Resistant Bacteria and Bacillus Cereus From
Libyan Journal of Medicine æ LETTER TO THE EDITOR Identification of multidrug-resistant bacteria and Bacillus cereus from healthcare workers and environmental surfaces in a hospital osocomial (hospital-acquired, healthcare-associated) Table 1. Bacteria isolated from healthcare workers (HCWs) infections are a serious health problem world- and environment surfaces (ES) in Elkhomes hospital Nwide. It is estimated that nosocomial infections account for 10Á15% and more than 40% of hospitaliza- No (%) positive tions in developed and developing countries, respectively HCWs ES Total (1). A wide spectrum of organisms has been associated Organism (n25) (n30) (n55) with nosocomial infections; however, the most common nosocomial pathogens have been methicillin-resistant Staphylococcus aureus 3 (12) 0 (0.0) 3 (5.5) Staphylococcus aureus (MRSA) and drug-resistant gram- S. haemolyticus 3 (12) 7 (23.3) 10 (18.2) negative bacteria (2). Outbreaks of nosocomial infections S. hominis 5 (20) 4 (13.3) 9 (16.4) initiated by colonized healthcare workers (HCWs) have S. epidermidis 3 (12) 1 (3.3) 4 (7.3) been reported previously (3). In addition, several studies S. cohnii subsp. cohnii 1 (4) 2 (6.7) 3 (5.5) suggest that contaminated environment surfaces (e.g. S. kloosii 1 (4) 2 (6.7) 3 (5.5) medical instruments) may play a role in the transmission S. saprophyticus 1 (4) 1 (3.3) 2 (3.6) of nosocomial pathogens (4, 5). S. capitis subsp. ureolyticus 1 (4) 1 (3.3) 2 (3.6) In the summer of 2013, premoistened sterile cotton- S. capitis subsp. capitis 2 (8) 0 (0.0) 2 (3.6) tipped swabs were used to collect specimens from the an- S. -

Note on Suspected Brown Recluse Spiders (Araneae: Sicariidae) in South Carolina
Faculty Research Note Note on Suspected Brown Recluse Spiders (Araneae: Sicariidae) in South Carolina Robert J. Wolff* South University, 9 Science Court, Columbia, SC 29203 The general public believes that brown recluse spiders (Loxosceles Filistatidae (Kukulcania hibernalis) 22 specimens reclusa) are widespread where they live and that these spiders are Lycosidae 21 (3 in one package, 5 in another) frequent causes of bites resulting in dermonecrosis. Research over the Pholcidae 17 past twenty years shows these reports to be unfounded. Vetter (2005) Miturgidae 8 examined 1,773 specimens sent in from across the U.S. as brown recluse Theridiidae 8 spiders and no specimens were found from areas outside the species Agelenidae 7 range, with the exception of a specimen from California. Araneidae 6 Clubionidae 6 The reported range of the brown recluse spider includes all or major Thomisidae 6 portions of Arkansas, Oklahoma, Texas, Louisiana, Alabama, Tennessee, Gnaphosidae 4 Kentucky, Illinois, Missouri and Kansas. Minor portions of the brown Corinnidae 3 recluse range were previously reported in Iowa, Indiana, Ohio, New Philodromidae 3 Mexico, North Carolina, Georgia, and South Carolina. The most recent Amaurobiidae 1 map (Vetter, 2015) does not include South Carolina, and only the far Pisauridae 1 western tip of North Carolina and northwestern corner of Georgia. Scytodidae (Scytodes thoracica) 1 Unidentifiable 4 Schuman and Caldwell (1991) found that South Carolina physicians reported treating 478 cases of brown recluse spider envenomations in 1990 alone. This seems like a very high number, unfortunately all or No brown recluses were identified from the specimens obtained in this almost all of these are probably not brown recluse spider bites.