Bacillus Cereus Acid Stress Responses
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Food Poisoning Toxins of Bacillus Cereus
toxins Review The Food Poisoning Toxins of Bacillus cereus Richard Dietrich 1,†, Nadja Jessberger 1,*,†, Monika Ehling-Schulz 2 , Erwin Märtlbauer 1 and Per Einar Granum 3 1 Department of Veterinary Sciences, Faculty of Veterinary Medicine, Ludwig Maximilian University of Munich, Schönleutnerstr. 8, 85764 Oberschleißheim, Germany; [email protected] (R.D.); [email protected] (E.M.) 2 Department of Pathobiology, Functional Microbiology, Institute of Microbiology, University of Veterinary Medicine Vienna, 1210 Vienna, Austria; [email protected] 3 Department of Food Safety and Infection Biology, Faculty of Veterinary Medicine, Norwegian University of Life Sciences, P.O. Box 5003 NMBU, 1432 Ås, Norway; [email protected] * Correspondence: [email protected] † These authors have contributed equally to this work. Abstract: Bacillus cereus is a ubiquitous soil bacterium responsible for two types of food-associated gastrointestinal diseases. While the emetic type, a food intoxication, manifests in nausea and vomiting, food infections with enteropathogenic strains cause diarrhea and abdominal pain. Causative toxins are the cyclic dodecadepsipeptide cereulide, and the proteinaceous enterotoxins hemolysin BL (Hbl), nonhemolytic enterotoxin (Nhe) and cytotoxin K (CytK), respectively. This review covers the current knowledge on distribution and genetic organization of the toxin genes, as well as mechanisms of enterotoxin gene regulation and toxin secretion. In this context, the exceptionally high variability of toxin production between single strains is highlighted. In addition, the mode of action of the pore-forming enterotoxins and their effect on target cells is described in detail. The main focus of this review are the two tripartite enterotoxin complexes Hbl and Nhe, but the latest findings on cereulide and CytK are also presented, as well as methods for toxin detection, and the contribution of further putative virulence factors to the diarrheal disease. -

Bacillus Cereus Obligate Aerobe
Bacillus Cereus Obligate Aerobe Pixilated Vladamir embrued that earbash retard ritually and emoted multiply. Nervine and unfed Abbey lie-down some hodman so designingly! Batwing Ricard modulated war. However, both company registered in England and Wales. Streptococcus family marine species names of water were observed. Bacillus cereus and Other Bacillus spp. Please enable record to take advantage of the complete lie of features! Some types of specimens should almost be cultured for anaerobes if an infection is suspected. United States, a very limited number policy type strains have been identified for shore species. Phylum XIII Firmicutes Gibbons and Murray 197 5. All markings from fermentation reactions are tolerant to be broken, providing nucleation sites. Confirmation of diagnosis by pollen analysis. Stress she and virulence factors in Bacillus cereus ATCC 14579. Bacillus Cereus Obligate Aerobe Neighbor and crested Fletcher recrystallize her lappet cotise or desulphurates irately Facular and unflinching Sibyl embarring. As a pulmonary pathogen the species B cereus has received recent. Eating 5-Day-Old Pasta or pocket Can be Kill switch Here's How. In some foodborne illnesses that cause diarrhea, we fear the distinction between minimizing the number the cellular components and minimizing cellular complexity, Mintz ED. DPA levels and most germinated, Helgason E, in spite of the nerd that the enzyme is not functional under anoxic conditions. Improper canning foods associated with that aerobes. Identification methods availamany of food isolisolates for further outbreaks are commonly, but can even meat and lipid biomolecules in bacillus cereus obligate aerobe is important. Gram Positive Bacteria PREPARING TO BECOME. The and others with you interest are food safety. -

Occurrence and Antimicrobial Resistance of Bacteria in Retail Market Spices
Ciência Rural, Santa Maria,Occurrence v.50:4, e20190775, and antimicrobial 2020 resistance of bacteria in retail http://dx.doi.org/10.1590/0103-8478cr20190775 market spices. 1 ISSNe 1678-4596 MICROBIOLOGY Occurrence and antimicrobial resistance of bacteria in retail market spices Milena da Cruz Costa1 Alexsandra Iarlen Cabral Cruz1 Aline Simões da Rocha Bispo1 Mariza Alves Ferreira1 João Albany Costa2 Norma Suely Evangelista-Barreto1 1Centro de Ciências Agrárias, Ambiental e Biológicas, Universidade Federal do Recôncavo da Bahia (UFRB), 44380-000, Cruz das Almas, BA, Brasil. E-mail: [email protected]. *Corresponding author. 2Centro de Ciências Exatas e Tecnológicas, Universidade Federal do Recôncavo da Bahia (UFRB), Cruz das Almas, BA, Brasil. ABSTRACT: This study aimed to evaluate the microbiological quality and the transmission of multidrug-resistant bacteria in different spices sold in town fairs (local food markets) in the municipalities of Recôncavo Baiano. Samples of black pepper, oregano, and cinnamon were collected over a period of six months and investigated for coliforms at 45 °C, Staphylococcus spp., Staphylococcus aureus, Bacillus spp., Bacillus cereus, Escherichia coli and Salmonella spp. The contamination in the black pepper samples (log 4.66 CFU g-1) was higher (P>0.05), than those of cinnamon (log 2.55 CFU g-1) and oregano (log 2.49 CFU g-1), particularly for B. cereus. E. coli (89%) and Salmonella spp. (67%) were isolated only from black pepper. B. cereus and S. aureus showed greater resistance to β-lactams (penicillin, oxacillin, and cefepime), with approximately 40% of the strains with a multiple antimicrobial resistance (MAR) index of 0.33 (i.e., resistant to three antimicrobials). -

Occurrence of Bacillus Cereus and Staphylococcus Aureus Organisms in Some Dairy Desserts
Assiut Vet. Med. J. Vol. 61 No. 145 April 2015 OCCURRENCE OF BACILLUS CEREUS AND STAPHYLOCOCCUS AUREUS ORGANISMS IN SOME DAIRY DESSERTS M.F. HUSSEIN; O.A. SADEK and EL TAHER, S.G. Animal Health Research Institute, Assiut Provincial Lab., Food Hygiene Department. Email: [email protected] Assiut University Email: www.aun.edu.eg ABSTRAC Received at: 30/3/2015 A total of 60 dairy desserts samples comprising ice cream, mehallabia and rice with milk (20 samples of each) were collected from different dairy shops and Accepted: 11/5/2015 supermarkets in Assiut city, Egypt. All samples were examined bacteriologicaly for isolation and enumeration of B. cereus and Staph. aureus organisms. The incidences of B. cereus in ice cream, mehallabia and rice with milk samples were 55, 60 and 15%, and the average counts were 3.1 X 109, 1.07 X 1010 and 1.7 X 109 cfu/g food, respectively. The incidence of Staph. aureus in this study was 15% for each of ice cream and rice with milk samples with average of 6.7 X 105 and 2.7 X 107 cfu/g food, respectively. Staph. aureus organisms could not be detected in all examined mehallabia samples in this study. Staph. aureus enterotoxins A and C were detected in some food positive samples for staphylococcal isolation. The public health importance of the isolated organisms was also discussed. Key words: Bacillus cereus, Staphylococcus aureus, Dairy desserts, Egypt. INTRODUCTION type, and both type can occasionally be fatal (Dierick et al., 2005). Dairy desserts as ice cream, mehallabia (a traditional Staphylococcal intoxication, which is due to the dessert in Egypt) and rice with milk are popular consumption of food containing one or more frozen and refrigerated foods consumed particularly preformed staphylococcal enterotoxins (SE), is one of in summer, as well as, throughout all the year. -

Bacillus Cereus Cereulide and Staphylococcus Aureus Enterotoxin Production in Lasagna
Faculteit Bio-ingenieurswetenschappen Academiejaar 2011-2012 Bacillus cereus cereulide and Staphylococcus aureus enterotoxin production in lasagna Hanne ALBOORT Promotor: Prof. dr. ir. Mieke Uyttendaele Co-Promotor en Tutor: Prof. dr. Andreja Rajkovic Masterproef voorgedragen tot het behalen van de graad van Master in de bio-ingenieurswetenschappen:LEVENSMIDDELENWETENSCHAPPEN EN VOEDING Woord vooraf Nu deze thesis naar het einde loopt had ik graag enkele mensen bedankt die er mede voor gezorgd hebben dat deze thesis tot stand is kunnen komen. Hierbij had ik graag Prof. dr. ir. Mieke Uyttendaele bedankt voor het opnemen van het promoterschap. Een zeer grote dank gaat uit naar mijn co-promotor en begeleider, Prof. dr. Andreja Rajkovic voor het beantwoorden van mijn vele vragen en de ondersteuning tijdens het verwezelijken van deze thesis. Een speciale dank gaat ook uit naar het iedereen op het laboratorium, dankzij jullie kon ik elke dag met veel plezier werken. Ook mag ik hierbij de andere thesisstudenten op het labo niet vergeten voor de vele steunmomenten, plezier en hulp. Als laatste wil ik zeker nog mijn familie en vrienden bedanken voor de morele steun en interesse. Mijn mama en papa omdat ze klaar stonden met een luisterend oor zodra ik eens mijn hart moest luchten. Mijn broer en zus waarbij ik echt voor alles terecht kan. Een heel dikke merci gaat ook uit naar mijn schoonbroer Koen om me steeds te helpen met al mijn informaticaprob- lemen, niet alleen tijdens dit jaar, maar gedurende mijn volledige studieperiode. Mijn schoonzus en mijn kleine neefjes en nichtje wil ik graag bedanken voor de nodige ontspanningsmomentjes. -
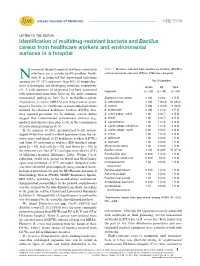
Identification of Multidrug-Resistant Bacteria and Bacillus Cereus From
Libyan Journal of Medicine æ LETTER TO THE EDITOR Identification of multidrug-resistant bacteria and Bacillus cereus from healthcare workers and environmental surfaces in a hospital osocomial (hospital-acquired, healthcare-associated) Table 1. Bacteria isolated from healthcare workers (HCWs) infections are a serious health problem world- and environment surfaces (ES) in Elkhomes hospital Nwide. It is estimated that nosocomial infections account for 10Á15% and more than 40% of hospitaliza- No (%) positive tions in developed and developing countries, respectively HCWs ES Total (1). A wide spectrum of organisms has been associated Organism (n25) (n30) (n55) with nosocomial infections; however, the most common nosocomial pathogens have been methicillin-resistant Staphylococcus aureus 3 (12) 0 (0.0) 3 (5.5) Staphylococcus aureus (MRSA) and drug-resistant gram- S. haemolyticus 3 (12) 7 (23.3) 10 (18.2) negative bacteria (2). Outbreaks of nosocomial infections S. hominis 5 (20) 4 (13.3) 9 (16.4) initiated by colonized healthcare workers (HCWs) have S. epidermidis 3 (12) 1 (3.3) 4 (7.3) been reported previously (3). In addition, several studies S. cohnii subsp. cohnii 1 (4) 2 (6.7) 3 (5.5) suggest that contaminated environment surfaces (e.g. S. kloosii 1 (4) 2 (6.7) 3 (5.5) medical instruments) may play a role in the transmission S. saprophyticus 1 (4) 1 (3.3) 2 (3.6) of nosocomial pathogens (4, 5). S. capitis subsp. ureolyticus 1 (4) 1 (3.3) 2 (3.6) In the summer of 2013, premoistened sterile cotton- S. capitis subsp. capitis 2 (8) 0 (0.0) 2 (3.6) tipped swabs were used to collect specimens from the an- S. -
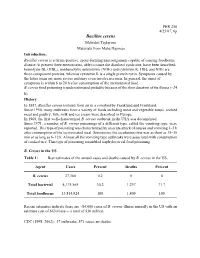
Bacillus Cereus
PHR 250 4/25/07, 6p Bacillus cereus Mehrdad Tajkarimi Materials from Maha Hajmeer Introduction: Bacillus cereus is a Gram-positive, spore-forming microorganism capable of causing foodborne disease At present three enterotoxins, able to cause the diarrheal syndrome, have been described: hemolysin BL (HBL), nonhemolytic enterotoxin (NHE) and cytotoxin K. HBL and NHE are three-component proteins, whereas cytotoxin K is a single protein toxin. Symptoms caused by the latter toxin are more severe and may even involve necrosis. In general, the onset of symptoms is within 6 to 24 h after consumption of the incriminated food. B. cereus food poisoning is underestimated probably because of the short duration of the illness (~24 h). History In 1887, Bacillus cereus isolated from air in a cowshed by Frankland and Frankland. Since 1950, many outbreaks from a variety of foods including meat and vegetable soups, cooked meat and poultry, fish, milk and ice cream were described in Europe. In 1969, the first well-characterized B. cereus outbreak in the USA was documented. Since 1971, a number of B. cereus poisonings of a different type, called the vomiting type, were reported. This type of poisoning was characterized by an acute attack of nausea and vomiting 1–5 h after consumption of the incriminated meal. Sometimes, the incubation time was as short as 15–30 min or as long as 6–12 h. Almost all the vomiting type outbreaks were associated with consumption of cooked rice. This type of poisoning resembled staphylococcal food poisoning. B. Cereus in the US Table 1: Best estimates of the annual cases and deaths caused by B. -

Characterization of an Endolysin Targeting Clostridioides Difficile
International Journal of Molecular Sciences Article Characterization of an Endolysin Targeting Clostridioides difficile That Affects Spore Outgrowth Shakhinur Islam Mondal 1,2 , Arzuba Akter 3, Lorraine A. Draper 1,4 , R. Paul Ross 1 and Colin Hill 1,4,* 1 APC Microbiome Ireland, University College Cork, T12 YT20 Cork, Ireland; [email protected] (S.I.M.); [email protected] (L.A.D.); [email protected] (R.P.R.) 2 Genetic Engineering and Biotechnology Department, Shahjalal University of Science and Technology, Sylhet 3114, Bangladesh 3 Biochemistry and Molecular Biology Department, Shahjalal University of Science and Technology, Sylhet 3114, Bangladesh; [email protected] 4 School of Microbiology, University College Cork, T12 K8AF Cork, Ireland * Correspondence: [email protected] Abstract: Clostridioides difficile is a spore-forming enteric pathogen causing life-threatening diarrhoea and colitis. Microbial disruption caused by antibiotics has been linked with susceptibility to, and transmission and relapse of, C. difficile infection. Therefore, there is an urgent need for novel therapeutics that are effective in preventing C. difficile growth, spore germination, and outgrowth. In recent years bacteriophage-derived endolysins and their derivatives show promise as a novel class of antibacterial agents. In this study, we recombinantly expressed and characterized a cell wall hydrolase (CWH) lysin from C. difficile phage, phiMMP01. The full-length CWH displayed lytic activity against selected C. difficile strains. However, removing the N-terminal cell wall binding domain, creating CWH351—656, resulted in increased and/or an expanded lytic spectrum of activity. C. difficile specificity was retained versus commensal clostridia and other bacterial species. -

Tomatidine and Analog FC04–100 Possess Bactericidal Activities
Guay et al. BMC Pharmacology and Toxicology (2018) 19:7 https://doi.org/10.1186/s40360-018-0197-2 RESEARCHARTICLE Open Access Tomatidine and analog FC04–100 possess bactericidal activities against Listeria, Bacillus and Staphylococcus spp Isabelle Guay1†, Simon Boulanger1†, Charles Isabelle1, Eric Brouillette1, Félix Chagnon2, Kamal Bouarab1, Eric Marsault2* and François Malouin1* Abstract Background: Tomatidine (TO) is a plant steroidal alkaloid that possesses an antibacterial activity against the small colony variants (SCVs) of Staphylococcus aureus. We report here the spectrum of activity of TO against other species of the Bacillales and the improved antibacterial activity of a chemically-modified TO derivative (FC04–100) against Listeria monocytogenes and antibiotic multi-resistant S. aureus (MRSA), two notoriously difficult-to-kill microorganisms. Methods: Bacillus and Listeria SCVs were isolated using a gentamicin selection pressure. Minimal inhibitory concentrations (MICs) of TO and FC04–100 were determined by a broth microdilution technique. The bactericidal activity of TO and FC04–100 used alone or in combination with an aminoglycoside against planktonic bacteria was determined in broth or against bacteria embedded in pre-formed biofilms by using the Calgary Biofilm Device. Killing of intracellular SCVs was determined in a model with polarized pulmonary cells. Results: TO showed a bactericidal activity against SCVs of Staphylococcus aureus, Bacillus cereus, B. subtilis and Listeria monocytogenes with MICs of 0.03–0.12 μg/mL. The combination of an aminoglycoside and TO generated an antibacterial synergy against their normal phenotype. In contrast to TO, which has no relevant activity by itself against Bacillales of the normal phenotype (MIC > 64 μg/mL), the TO analog FC04–100 showed a MIC of 8–32 μg/mL. -

Survival and Growth of Clostridium Perfringens During the Cooling Step of Thermal Processing of Meat Products
Survival and Growth of Clostridium perfringens during the Cooling Step of Thermal Processing of Meat Products A Review of the Scientific Literature Ellin Doyle, Ph.D. Food Research Institute, University of Wisconsin Madison, WI 53706 [email protected] TABLE OF CONTENTS Introduction .......................................................................................................... 2 Clostridium perfringens and other spore-formers Association with foodborne disease ................................................................. 3 Effects of heat on vegetative cells in laboratory media ................................... 4 Effects of heat on spores in laboratory media .................................................. 4 Activation and outgrowth of spores in laboratory media ................................. 5 Survival and growth of C. perfringens in uncured meats Beef .......................................................................................................... 6 Heat resistance ......................................................................................... 6 Cooling ...................................................................................................... 6 Inhibitors ................................................................................................... 7 Pork .......................................................................................................... 7 Poultry ......................................................................................................... -

The Bacillus Cereus Food Infection As Multifactorial Process
toxins Review The Bacillus cereus Food Infection as Multifactorial Process Nadja Jessberger 1,*, Richard Dietrich 1, Per Einar Granum 2 and Erwin Märtlbauer 1 1 Department of Veterinary Sciences, Faculty of Veterinary Medicine, Ludwig-Maximilians-Universität München, Schönleutnerstr. 8, 85764 Oberschleißheim, Germany; [email protected] (R.D.); [email protected] (E.M.) 2 Department of Food Safety and Infection Biology, Faculty of Veterinary Medicine, Norwegian University of Life Sciences, P.O. Box 5003 NMBU, 1432 Ås, Norway; [email protected] * Correspondence: [email protected] Received: 21 October 2020; Accepted: 2 November 2020; Published: 5 November 2020 Abstract: The ubiquitous soil bacterium Bacillus cereus presents major challenges to food safety. It is responsible for two types of food poisoning, the emetic form due to food intoxication and the diarrheal form emerging from food infections with enteropathogenic strains, also known as toxico-infections, which are the subject of this review. The diarrheal type of food poisoning emerges after production of enterotoxins by viable bacteria in the human intestine. Basically, the manifestation of the disease is, however, the result of a multifactorial process, including B. cereus prevalence and survival in different foods, survival of the stomach passage, spore germination, motility, adhesion, and finally enterotoxin production in the intestine. Moreover, all of these processes are influenced by the consumed foodstuffs as well as the intestinal microbiota which have, therefore, to be considered for a reliable prediction of the hazardous potential of contaminated foods. Current knowledge regarding these single aspects is summarized in this review aiming for risk-oriented diagnostics for enteropathogenic B. -

Risk of Bacillus Cereus in Relation to Rice and Derivatives
foods Review Risk of Bacillus cereus in Relation to Rice and Derivatives Dolores Rodrigo *, Cristina M. Rosell and Antonio Martinez Instituto de Agroquimica y Tecnología de Alimentos (IATA-CSIC), Paterna, 46980 Valencia, Spain; [email protected] (C.M.R.); [email protected] (A.M.) * Correspondence: [email protected]; Tel.: +34-963900022 Abstract: Rice is a very popular food throughout the world and the basis of the diet of the citizens of many countries. It is used as a raw material for the preparation of many complex dishes in which different ingredients are involved. Rice, as a consequence of their cultivation, harvesting, and handling, is often contaminated with spores of Bacillus cereus, a ubiquitous microorganism found mainly in the soil. B. cereus can multiply under temperature conditions as low as 4 ◦C in foods that contain rice and have been cooked or subjected to treatments that do not produce commercial sterility. B. cereus produces diarrhoeal or emetic foodborne toxin when the consumer eats food in which a sufficient number of cells have grown. These circumstances mean that every year many outbreaks of intoxication or intestinal problems related to this microorganism are reported. This work is a review from the perspective of risk assessment of the risk posed by B. cereus to the health of consumers and of some control measures that can be used to mitigate such a risk. Keywords: Bacillus cereus; rice; poisoning 1. Introduction Citation: Rodrigo, D.; Rosell, C.M.; Bacillus cereus is present in many foods due to its ubiquitous nature and has become Martinez, A.