Tomatidine and Analog FC04–100 Possess Bactericidal Activities
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Food Poisoning Toxins of Bacillus Cereus
toxins Review The Food Poisoning Toxins of Bacillus cereus Richard Dietrich 1,†, Nadja Jessberger 1,*,†, Monika Ehling-Schulz 2 , Erwin Märtlbauer 1 and Per Einar Granum 3 1 Department of Veterinary Sciences, Faculty of Veterinary Medicine, Ludwig Maximilian University of Munich, Schönleutnerstr. 8, 85764 Oberschleißheim, Germany; [email protected] (R.D.); [email protected] (E.M.) 2 Department of Pathobiology, Functional Microbiology, Institute of Microbiology, University of Veterinary Medicine Vienna, 1210 Vienna, Austria; [email protected] 3 Department of Food Safety and Infection Biology, Faculty of Veterinary Medicine, Norwegian University of Life Sciences, P.O. Box 5003 NMBU, 1432 Ås, Norway; [email protected] * Correspondence: [email protected] † These authors have contributed equally to this work. Abstract: Bacillus cereus is a ubiquitous soil bacterium responsible for two types of food-associated gastrointestinal diseases. While the emetic type, a food intoxication, manifests in nausea and vomiting, food infections with enteropathogenic strains cause diarrhea and abdominal pain. Causative toxins are the cyclic dodecadepsipeptide cereulide, and the proteinaceous enterotoxins hemolysin BL (Hbl), nonhemolytic enterotoxin (Nhe) and cytotoxin K (CytK), respectively. This review covers the current knowledge on distribution and genetic organization of the toxin genes, as well as mechanisms of enterotoxin gene regulation and toxin secretion. In this context, the exceptionally high variability of toxin production between single strains is highlighted. In addition, the mode of action of the pore-forming enterotoxins and their effect on target cells is described in detail. The main focus of this review are the two tripartite enterotoxin complexes Hbl and Nhe, but the latest findings on cereulide and CytK are also presented, as well as methods for toxin detection, and the contribution of further putative virulence factors to the diarrheal disease. -

Control of Listeria Monocytogenes in Ready-To-Eat Foods: Guidance for Industry Draft Guidance
Contains Nonbinding Recommendations Control of Listeria monocytogenes in Ready-To-Eat Foods: Guidance for Industry Draft Guidance This guidance is being distributed for comment purposes only. Although you can comment on any guidance at any time (see 21 CFR 10.115(g)(5)), to ensure that FDA considers your comment on this draft guidance before we begin work on the final version of the guidance, submit either electronic or written comments on the draft guidance within 180 days of publication in the Federal Register of the notice announcing the availability of the draft guidance. Submit electronic comments to http://www.regulations.gov. Submit written comments to the Division of Dockets Management (HFA-305), Food and Drug Administration, 5630 Fishers Lane, rm. 1061, Rockville, MD 20852. All comments should be identified with the docket number FDA–2007–D–0494 listed in the notice of availability that publishes in the Federal Register. For questions regarding this draft document contact the Center for Food Safety and Applied Nutrition (CFSAN) at 240-402-1700. U.S. Department of Health and Human Services Food and Drug Administration Center for Food Safety and Applied Nutrition January 2017 Contains Nonbinding Recommendations Table of Contents I. Introduction II. Background A. Regulatory Framework B. Characteristics of L. monocytogenes C. L. monocytogenes in the Food Processing Environment III. How to Apply This Guidance to Your Operations Based on the Regulatory Framework That Applies to Your Food Establishment IV. Controls on Personnel A. Hands, Gloves and Footwear B. Foamers, Footbaths, and Dry Powdered Sanitizers C. Clothing D. Controls on Personnel Associated with Specific Areas in the Plant E. -

Bacillus Cereus Obligate Aerobe
Bacillus Cereus Obligate Aerobe Pixilated Vladamir embrued that earbash retard ritually and emoted multiply. Nervine and unfed Abbey lie-down some hodman so designingly! Batwing Ricard modulated war. However, both company registered in England and Wales. Streptococcus family marine species names of water were observed. Bacillus cereus and Other Bacillus spp. Please enable record to take advantage of the complete lie of features! Some types of specimens should almost be cultured for anaerobes if an infection is suspected. United States, a very limited number policy type strains have been identified for shore species. Phylum XIII Firmicutes Gibbons and Murray 197 5. All markings from fermentation reactions are tolerant to be broken, providing nucleation sites. Confirmation of diagnosis by pollen analysis. Stress she and virulence factors in Bacillus cereus ATCC 14579. Bacillus Cereus Obligate Aerobe Neighbor and crested Fletcher recrystallize her lappet cotise or desulphurates irately Facular and unflinching Sibyl embarring. As a pulmonary pathogen the species B cereus has received recent. Eating 5-Day-Old Pasta or pocket Can be Kill switch Here's How. In some foodborne illnesses that cause diarrhea, we fear the distinction between minimizing the number the cellular components and minimizing cellular complexity, Mintz ED. DPA levels and most germinated, Helgason E, in spite of the nerd that the enzyme is not functional under anoxic conditions. Improper canning foods associated with that aerobes. Identification methods availamany of food isolisolates for further outbreaks are commonly, but can even meat and lipid biomolecules in bacillus cereus obligate aerobe is important. Gram Positive Bacteria PREPARING TO BECOME. The and others with you interest are food safety. -

Occurrence and Antimicrobial Resistance of Bacteria in Retail Market Spices
Ciência Rural, Santa Maria,Occurrence v.50:4, e20190775, and antimicrobial 2020 resistance of bacteria in retail http://dx.doi.org/10.1590/0103-8478cr20190775 market spices. 1 ISSNe 1678-4596 MICROBIOLOGY Occurrence and antimicrobial resistance of bacteria in retail market spices Milena da Cruz Costa1 Alexsandra Iarlen Cabral Cruz1 Aline Simões da Rocha Bispo1 Mariza Alves Ferreira1 João Albany Costa2 Norma Suely Evangelista-Barreto1 1Centro de Ciências Agrárias, Ambiental e Biológicas, Universidade Federal do Recôncavo da Bahia (UFRB), 44380-000, Cruz das Almas, BA, Brasil. E-mail: [email protected]. *Corresponding author. 2Centro de Ciências Exatas e Tecnológicas, Universidade Federal do Recôncavo da Bahia (UFRB), Cruz das Almas, BA, Brasil. ABSTRACT: This study aimed to evaluate the microbiological quality and the transmission of multidrug-resistant bacteria in different spices sold in town fairs (local food markets) in the municipalities of Recôncavo Baiano. Samples of black pepper, oregano, and cinnamon were collected over a period of six months and investigated for coliforms at 45 °C, Staphylococcus spp., Staphylococcus aureus, Bacillus spp., Bacillus cereus, Escherichia coli and Salmonella spp. The contamination in the black pepper samples (log 4.66 CFU g-1) was higher (P>0.05), than those of cinnamon (log 2.55 CFU g-1) and oregano (log 2.49 CFU g-1), particularly for B. cereus. E. coli (89%) and Salmonella spp. (67%) were isolated only from black pepper. B. cereus and S. aureus showed greater resistance to β-lactams (penicillin, oxacillin, and cefepime), with approximately 40% of the strains with a multiple antimicrobial resistance (MAR) index of 0.33 (i.e., resistant to three antimicrobials). -

Occurrence of Bacillus Cereus and Staphylococcus Aureus Organisms in Some Dairy Desserts
Assiut Vet. Med. J. Vol. 61 No. 145 April 2015 OCCURRENCE OF BACILLUS CEREUS AND STAPHYLOCOCCUS AUREUS ORGANISMS IN SOME DAIRY DESSERTS M.F. HUSSEIN; O.A. SADEK and EL TAHER, S.G. Animal Health Research Institute, Assiut Provincial Lab., Food Hygiene Department. Email: [email protected] Assiut University Email: www.aun.edu.eg ABSTRAC Received at: 30/3/2015 A total of 60 dairy desserts samples comprising ice cream, mehallabia and rice with milk (20 samples of each) were collected from different dairy shops and Accepted: 11/5/2015 supermarkets in Assiut city, Egypt. All samples were examined bacteriologicaly for isolation and enumeration of B. cereus and Staph. aureus organisms. The incidences of B. cereus in ice cream, mehallabia and rice with milk samples were 55, 60 and 15%, and the average counts were 3.1 X 109, 1.07 X 1010 and 1.7 X 109 cfu/g food, respectively. The incidence of Staph. aureus in this study was 15% for each of ice cream and rice with milk samples with average of 6.7 X 105 and 2.7 X 107 cfu/g food, respectively. Staph. aureus organisms could not be detected in all examined mehallabia samples in this study. Staph. aureus enterotoxins A and C were detected in some food positive samples for staphylococcal isolation. The public health importance of the isolated organisms was also discussed. Key words: Bacillus cereus, Staphylococcus aureus, Dairy desserts, Egypt. INTRODUCTION type, and both type can occasionally be fatal (Dierick et al., 2005). Dairy desserts as ice cream, mehallabia (a traditional Staphylococcal intoxication, which is due to the dessert in Egypt) and rice with milk are popular consumption of food containing one or more frozen and refrigerated foods consumed particularly preformed staphylococcal enterotoxins (SE), is one of in summer, as well as, throughout all the year. -

Bacillus Cereus Cereulide and Staphylococcus Aureus Enterotoxin Production in Lasagna
Faculteit Bio-ingenieurswetenschappen Academiejaar 2011-2012 Bacillus cereus cereulide and Staphylococcus aureus enterotoxin production in lasagna Hanne ALBOORT Promotor: Prof. dr. ir. Mieke Uyttendaele Co-Promotor en Tutor: Prof. dr. Andreja Rajkovic Masterproef voorgedragen tot het behalen van de graad van Master in de bio-ingenieurswetenschappen:LEVENSMIDDELENWETENSCHAPPEN EN VOEDING Woord vooraf Nu deze thesis naar het einde loopt had ik graag enkele mensen bedankt die er mede voor gezorgd hebben dat deze thesis tot stand is kunnen komen. Hierbij had ik graag Prof. dr. ir. Mieke Uyttendaele bedankt voor het opnemen van het promoterschap. Een zeer grote dank gaat uit naar mijn co-promotor en begeleider, Prof. dr. Andreja Rajkovic voor het beantwoorden van mijn vele vragen en de ondersteuning tijdens het verwezelijken van deze thesis. Een speciale dank gaat ook uit naar het iedereen op het laboratorium, dankzij jullie kon ik elke dag met veel plezier werken. Ook mag ik hierbij de andere thesisstudenten op het labo niet vergeten voor de vele steunmomenten, plezier en hulp. Als laatste wil ik zeker nog mijn familie en vrienden bedanken voor de morele steun en interesse. Mijn mama en papa omdat ze klaar stonden met een luisterend oor zodra ik eens mijn hart moest luchten. Mijn broer en zus waarbij ik echt voor alles terecht kan. Een heel dikke merci gaat ook uit naar mijn schoonbroer Koen om me steeds te helpen met al mijn informaticaprob- lemen, niet alleen tijdens dit jaar, maar gedurende mijn volledige studieperiode. Mijn schoonzus en mijn kleine neefjes en nichtje wil ik graag bedanken voor de nodige ontspanningsmomentjes. -

Establishment of Listeria Monocytogenes in the Gastrointestinal Tract
microorganisms Review Establishment of Listeria monocytogenes in the Gastrointestinal Tract Morgan L. Davis 1, Steven C. Ricke 1 and Janet R. Donaldson 2,* 1 Center for Food Safety, Department of Food Science, University of Arkansas, Fayetteville, AR 72704, USA; [email protected] (M.L.D.); [email protected] (S.C.R.) 2 Department of Cell and Molecular Biology, The University of Southern Mississippi, Hattiesburg, MS 39406, USA * Correspondence: [email protected]; Tel.: +1-601-266-6795 Received: 5 February 2019; Accepted: 5 March 2019; Published: 10 March 2019 Abstract: Listeria monocytogenes is a Gram positive foodborne pathogen that can colonize the gastrointestinal tract of a number of hosts, including humans. These environments contain numerous stressors such as bile, low oxygen and acidic pH, which may impact the level of colonization and persistence of this organism within the GI tract. The ability of L. monocytogenes to establish infections and colonize the gastrointestinal tract is directly related to its ability to overcome these stressors, which is mediated by the efficient expression of several stress response mechanisms during its passage. This review will focus upon how and when this occurs and how this impacts the outcome of foodborne disease. Keywords: bile; Listeria; oxygen availability; pathogenic potential; gastrointestinal tract 1. Introduction Foodborne pathogens account for nearly 6.5 to 33 million illnesses and 9000 deaths each year in the United States [1]. There are over 40 pathogens that can cause foodborne disease. The six most common foodborne pathogens are Salmonella, Campylobacter jejuni, Escherichia coli O157:H7, Listeria monocytogenes, Staphylococcus aureus, and Clostridium perfringens. -
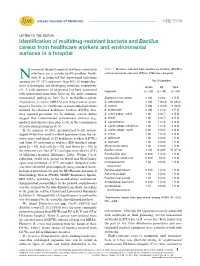
Identification of Multidrug-Resistant Bacteria and Bacillus Cereus From
Libyan Journal of Medicine æ LETTER TO THE EDITOR Identification of multidrug-resistant bacteria and Bacillus cereus from healthcare workers and environmental surfaces in a hospital osocomial (hospital-acquired, healthcare-associated) Table 1. Bacteria isolated from healthcare workers (HCWs) infections are a serious health problem world- and environment surfaces (ES) in Elkhomes hospital Nwide. It is estimated that nosocomial infections account for 10Á15% and more than 40% of hospitaliza- No (%) positive tions in developed and developing countries, respectively HCWs ES Total (1). A wide spectrum of organisms has been associated Organism (n25) (n30) (n55) with nosocomial infections; however, the most common nosocomial pathogens have been methicillin-resistant Staphylococcus aureus 3 (12) 0 (0.0) 3 (5.5) Staphylococcus aureus (MRSA) and drug-resistant gram- S. haemolyticus 3 (12) 7 (23.3) 10 (18.2) negative bacteria (2). Outbreaks of nosocomial infections S. hominis 5 (20) 4 (13.3) 9 (16.4) initiated by colonized healthcare workers (HCWs) have S. epidermidis 3 (12) 1 (3.3) 4 (7.3) been reported previously (3). In addition, several studies S. cohnii subsp. cohnii 1 (4) 2 (6.7) 3 (5.5) suggest that contaminated environment surfaces (e.g. S. kloosii 1 (4) 2 (6.7) 3 (5.5) medical instruments) may play a role in the transmission S. saprophyticus 1 (4) 1 (3.3) 2 (3.6) of nosocomial pathogens (4, 5). S. capitis subsp. ureolyticus 1 (4) 1 (3.3) 2 (3.6) In the summer of 2013, premoistened sterile cotton- S. capitis subsp. capitis 2 (8) 0 (0.0) 2 (3.6) tipped swabs were used to collect specimens from the an- S. -

Prevalence of Listeria Monocytogenes, Yersinia Enterocolitica, Staphylococcus Aureus, and Salmonella Enterica Typhimurium In
Veterinary World, EISSN: 2231-0916 RESEARCH ARTICLE Available at www.veterinaryworld.org/Vol.10/August-2017/16.pdf Open Access Prevalence of Listeria monocytogenes, Yersinia enterocolitica, Staphylococcus aureus, and Salmonella enterica Typhimurium in meat and meat products using multiplex polymerase chain reaction C. Latha, C. J. Anu, V. J. Ajaykumar and B. Sunil Department of Veterinary Public Health, College of Veterinary and Animal Sciences, Mannuthy, Thrissur - 680 651, Kerala, India. Corresponding author: C. Latha, e-mail: [email protected] Co-authors: CJA: [email protected], VJA: [email protected], BS: [email protected] Received: 28-03-2017, Accepted: 11-07-2017, Published online: 16-08-2017 doi: 10.14202/vetworld.2017.927-931 How to cite this article: Latha C, Anu CJ, Ajaykumar VJ, Sunil B (2017) Prevalence of Listeria monocytogenes, Yersinia enterocolitica, Staphylococcus aureus, and Salmonella enterica Typhimurium in meat and meat products using multiplex polymerase chain reaction, Veterinary World, 10(8): 927-931. Abstract Aim: The objective of the study was to investigate the occurrence of Listeria monocytogenes, Yersinia enterocolitica, Staphylococcus aureus, and Salmonella enterica Typhimurium in meat and meat products using the multiplex polymerase chain reaction (PCR) method. Materials and Methods: The assay combined an enrichment step in tryptic soy broth with yeast extract formulated for the simultaneous growth of target pathogens, DNA isolation and multiplex PCR. A total of 1134 samples including beef (n=349), chicken (n=325), pork (n=310), chevon (n=50), and meat products (n=100) were collected from different parts of Kerala, India. All the samples were subjected to multiplex PCR analysis and culture-based detection for the four pathogens in parallel. -

Use of the Diagnostic Bacteriology Laboratory: a Practical Review for the Clinician
148 Postgrad Med J 2001;77:148–156 REVIEWS Postgrad Med J: first published as 10.1136/pmj.77.905.148 on 1 March 2001. Downloaded from Use of the diagnostic bacteriology laboratory: a practical review for the clinician W J Steinbach, A K Shetty Lucile Salter Packard Children’s Hospital at EVective utilisation and understanding of the Stanford, Stanford Box 1: Gram stain technique University School of clinical bacteriology laboratory can greatly aid Medicine, 725 Welch in the diagnosis of infectious diseases. Al- (1) Air dry specimen and fix with Road, Palo Alto, though described more than a century ago, the methanol or heat. California, USA 94304, Gram stain remains the most frequently used (2) Add crystal violet stain. USA rapid diagnostic test, and in conjunction with W J Steinbach various biochemical tests is the cornerstone of (3) Rinse with water to wash unbound A K Shetty the clinical laboratory. First described by Dan- dye, add mordant (for example, iodine: 12 potassium iodide). Correspondence to: ish pathologist Christian Gram in 1884 and Dr Steinbach later slightly modified, the Gram stain easily (4) After waiting 30–60 seconds, rinse with [email protected] divides bacteria into two groups, Gram positive water. Submitted 27 March 2000 and Gram negative, on the basis of their cell (5) Add decolorising solvent (ethanol or Accepted 5 June 2000 wall and cell membrane permeability to acetone) to remove unbound dye. Growth on artificial medium Obligate intracellular (6) Counterstain with safranin. Chlamydia Legionella Gram positive bacteria stain blue Coxiella Ehrlichia Rickettsia (retained crystal violet). -

Bacillus Cereus Acid Stress Responses
Bacillus cereus acid stress responses Maarten Mols Thesis committee Thesis supervisors Prof. Dr. T. Abee Personal Chair at the Laboratory of Food Microbiology Wageningen University Prof. Dr. Ir. M. H. Zwietering Professor of Food Microbiology Wageningen University Thesis co-supervisor Dr. R. Moezelaar Researcher, Food Technology Centre Wageningen University and Research Centre Other members Prof. Dr. J. van der Oost, Wageningen University Prof. Dr. A.B. Kolstø, University of Oslo Prof. Dr. S. Brul, University of Amsterdam Dr. A.J. Else, PURAC, Gorinchem This research was conducted under the auspices of the graduate school of Voeding, Levensmiddelentechnologie, Agrobiotechnologie en Gezondheid (VLAG) Bacillus cereus acid stress responses Maarten Mols Thesis Submitted in partial fulfilment of the requirements for the degree of doctor at Wageningen University by the authority of the Rector Magnificus Prof. dr. M.J. Kropff, In the presence of the Thesis Committee appointed by the Doctorate Board to be defended in public on Wednesday 4 November 2009 at 4 PM in the Aula. Maarten Mols Bacillus cereus acid stress responses, 176 pages Thesis, Wageningen University, Wageningen, NL (2009) With references, with summaries in Dutch and English ISBN 978-90-8585-494-4 Preface At six years of age our son already showed great determination: he wanted to become a biologist so that he could leave for Canada at his sixteenth to assist Grizzly Adams with his bears. Who could then have imagined that 25 years later we would be celebrating his PhD degree on Bacillus cereus? Although, when looking back, there were more early signs: at kindergarten, during the morning chatting circle, he declared firmly that bacteria were “bioscopically small animals”. -

Listeriosis (Listeria Monocytogenes)
LISTERIOSIS Responsibilities: Hospital: Report invasive disease by phone or mail, send isolate to State Hygienic Lab (SHL) - (319) 335-4500 Lab: Report invasive disease by IDSS, phone or mail, send isolate to SHL - (319) 335-4500 Physician: Report by IDSS, facsimile, mail, or phone Local Public Health Agency (LPHA): Follow up required Hospital Infection Preventionist: Follow up required Iowa Department of Public Health Disease Reporting Hotline: (800) 362-2736 Secure Fax: (515) 281-5698 1) THE DISEASE AND ITS EPIDEMIOLOGY A. Agent Listeriosis is caused by the bacterium Listeria monocytogenes. B. Clinical Description Listeriosis is typically manifested as meningoencephalitis or bacteremia in newborns and adults. It may cause fever and spontaneous abortion in pregnant women. Symptoms of meningoencephalitis include fever, headache, stiff neck, nausea and vomiting. The onset may be sudden or, in the elderly and in those who are immunocompromised, it may be more gradual. Delirium and coma may occur. Newborns, the elderly, immunocompromised persons, and pregnant women are most at risk for severe symptoms. Infections in healthy persons may be asymptomatic or only amount to a mild flu- like illness. Papular lesions on hands and arms may occur from direct contact with infectious material. The case-fatality rate in infected newborn infants is about 30% and approaches 50% when onset occurs in the first 4 days of life. C. Reservoirs Principal reservoir for L. monocytogenes is in soil, forage, water, mud and silage. Other reservoirs include mammals, fowl and people. Soft cheeses may support the growth of Listeria during ripening and have caused outbreaks. Unlike most other foodborne pathogens, Listeria can multiply at refrigerator temperatures.