Small-Scale Analysis of Phytoecdysteroids in Seeds by HPLC- DAD-MS for the Identification and Quantification of Specific Analogues, Dereplication and Chemotaxonomy
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Botanical Survey of Joseph Quer's Flora Española
A botanical survey of Joseph Quer's Flora española Author(s): Carlos Aedo, Marta Fernández-Albert, Patricia Barberá, Antoni Buira, Alejandro Quintanar, Leopoldo Medina & Ramón Morales Source: Willdenowia, 47(3):243-258. Published By: Botanic Garden and Botanical Museum Berlin (BGBM) https://doi.org/10.3372/wi.47.47308 URL: http://www.bioone.org/doi/full/10.3372/wi.47.47308 BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use. Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder. BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research. Willdenowia Annals of the Botanic Garden and Botanical Museum Berlin-Dahlem CARLOS AEDO1*, MARTA FERNÁNDEZ-ALBERT1, PATRICIA BARBERÁ1, ANTONI BUIRA1, ALEJANDRO QUINTANAR1, LEOPOLDO MEDINA1 & RAMÓN MORALES1 A botanical survey of Joseph Quer’s Flora española Version of record first published online on 15 November 2017 ahead of inclusion in December 2017 issue. Abstract: We examine various aspects of Joseph Quer’s Flora española (1762 – 1764, 1784), taking into considera- tion the contributions made by Casimiro Gómez Ortega and proposing that he be credited as a co-author on the last two volumes of the work. -

Systematic Studies of the South African Campanulaceae Sensu Stricto with an Emphasis on Generic Delimitations
Town The copyright of this thesis rests with the University of Cape Town. No quotation from it or information derivedCape from it is to be published without full acknowledgement of theof source. The thesis is to be used for private study or non-commercial research purposes only. University Systematic studies of the South African Campanulaceae sensu stricto with an emphasis on generic delimitations Christopher Nelson Cupido Thesis presented for the degree of DOCTOR OF PHILOSOPHY in the Department of Botany Town UNIVERSITY OF CAPECape TOWN of September 2009 University Roella incurva Merciera eckloniana Microcodon glomeratus Prismatocarpus diffusus Town Wahlenbergia rubioides Cape of Wahlenbergia paniculata (blue), W. annularis (white) Siphocodon spartioides University Rhigiophyllum squarrosum Wahlenbergia procumbens Representatives of Campanulaceae diversity in South Africa ii Town Dedicated to Ursula, Denroy, Danielle and my parents Cape of University iii Town DECLARATION Cape I confirm that this is my ownof work and the use of all material from other sources has been properly and fully acknowledged. University Christopher N Cupido Cape Town, September 2009 iv Systematic studies of the South African Campanulaceae sensu stricto with an emphasis on generic delimitations Christopher Nelson Cupido September 2009 ABSTRACT The South African Campanulaceae sensu stricto, comprising 10 genera, represent the most diverse lineage of the family in the southern hemisphere. In this study two phylogenies are reconstructed using parsimony and Bayesian methods. A family-level phylogeny was estimated to test the monophyly and time of divergence of the South African lineage. This analysis, based on a published ITS phylogeny and an additional ten South African taxa, showed a strongly supported South African clade sister to the campanuloids. -

Cytotoxic Effect of Some Digitalis Species; a Study of Selectivity
FABAD J. Pharm. Sci., 43(1), 25-29, 2018 RESEARCH ARTICLE Cytotoxic Effect of Some Digitalis Species; A Study of Selectivity Vahap Murat KUTLUAY *°, İclal SARAÇOĞLU* Cytotoxic Effect of Some Digitalis Species; A Study of Bazı Digitalis Türlerinin Sitotoksik Etkisi; Bir Selektivite Selectivity Çalışması SUMMARY ÖZET Our previous research, on the aerial parts and roots of three Daha önceki çalışmamızda üç farklı Digitalis türünün (D.davisiana different Digitalis species; D.davisiana Heywood, D.viridiflora Heywood, D.viridiflora Lindley ve D.grandiflora Miller) toprak üstü Lindley and D.grandiflora Miller against HEp-2 and HepG2 kısımlarının ve köklerinin HEp-2 ve HepG2 kanser hücre serileri üzerinde, cancer cell lines showed that aerial parts were more cytotoxic than toprak üstü kısımların köklere göre daha sitotoksik olduğu gösterilmiştir. roots. So for further research a comparative cytotoxic activity study Bu nedenle D.davisiana, D.viridiflora ve D.grandiflora’nın toprak üstü was performed on the aerial parts of D.davisiana, D.viridiflora kısımların üzerinde gerçekleştirilen araştırma ilerletilerek HeLa kanser and D.grandiflora against HeLa cancer cell line. 3Y1 non- hücresi üzerindeki sitotoksik etkisi incelenmiştir. Sağlıklı bir hücre cancerous cell line was used to determine the selectivity. All three olan 3Y1, selektivitenin değerlendirilebilmesi için kullanılmıştır. Her üç ekstre de HeLa hücreleri üzerinde kuvvetli sitotoksik etki göstermiş extracts showed strong cytotoxic activity on HeLa cells with IC50 values less than 15 μg/ml. However, observed cytotoxic activity on ve IC50 değerleri 15 μg/ml’nin altında hesaplanmıştır. Ekstrelerin 3Y1 hücreleri üzerinde sitotoksik etkileri daha düşük olmakla birlikte 3Y1 cells was found to be lower than that of HeLa cells with IC50 values in a range of 94.7-772.3 μg/ml. -

Hardy Herbaceous & Alpine Plants : Wholesale Catalog, Spring 1928
Historic, Archive Document Do not assume content reflects current scientific knowledge, policies, or practices. 5^ IIIBP A I n«f«nrin|| l * FEB 2 5 $2r -* Hardy Herbaceous 6-' Alpine Plants i:w REG'D U. S. PAT. OFF. 9fce WAYSIDE GARDENS CO. Mentor. Ohio Wholesale Catalog Spring 1928 N presenting this catalog of HARDY PLANTS and ROCK PLANTS, we do so with the kindest regards to you who have favored us heretofore with your orders, and by continuing the To the excellence of our products and efficiency of serv¬ ice, we hope to merit your future patronage. Trade From those unacquainted with us we solicit a trial, and feel assured that the quality of our plants will prove to meet your most exacting demands. No order is too large for us to handle or too small to receive careful attention. Quality of plants and attention to every detail you entrust to us is assured. We have spared neither trouble nor expense in the production of the plants enumerated in this catalog and we offer them with the fullest con¬ fidence as to their general excellence, reliability, and ability to create a perfect garden within an exceptionally short space of time. Terms of business will be found on page 79, and a complete index to this catalog on page 80, the use of which will facilitate the study of its contents. The Wayside Gardens Co. Mentor, Ohio February first, 1928 E. H. SCHULTZ, President J. J. GRULLEMANS, Sec’y-Treas. Cardin cm ; k VV YORK CHICAGO BOSTON TORONTO SANTA BARBARA THE WORLD 8 WORK COUNTRY LIFE «Advertising CARDEN * HQME BUILDER ThECOUNTRYLIFE-PRESS ‘"Department RADIO BROADCAST GARDEN-CITY'"NEW-YORK. -

Taxonomic Composition and Ecological Characteristics of the Endemic Flora of the Lower Duero Basin (Iberian Peninsula)
Acta Botanica Gallica ISSN: 1253-8078 (Print) 2166-3408 (Online) Journal homepage: http://www.tandfonline.com/loi/tabg20 Taxonomic composition and ecological characteristics of the endemic flora of the lower Duero Basin (Iberian Peninsula) Francisco Amich , Sonia Bernardos , Carlos Aguiar , Javier Fernández-Diez & Antonio Crespí To cite this article: Francisco Amich , Sonia Bernardos , Carlos Aguiar , Javier Fernández-Diez & Antonio Crespí (2004) Taxonomic composition and ecological characteristics of the endemic flora of the lower Duero Basin (Iberian Peninsula), Acta Botanica Gallica, 151:4, 341-352, DOI: 10.1080/12538078.2004.10515439 To link to this article: http://dx.doi.org/10.1080/12538078.2004.10515439 Published online: 27 Apr 2013. Submit your article to this journal Article views: 104 View related articles Citing articles: 7 View citing articles Full Terms & Conditions of access and use can be found at http://www.tandfonline.com/action/journalInformation?journalCode=tabg20 Download by: [b-on: Biblioteca do conhecimento online IPB] Date: 24 July 2017, At: 04:09 Acta Bot. Gallica, 2004, 15/ (4), 341-352. Taxonomic composition and ecological characteristics of the endemic flora of the lower Duero Basin (Iberian Peninsula) by Francisco Amiche), Sonia Bemardose), Carlos Aguiare), Javier Femandez-Diez(1) and Antonio Crespi e) (/) Department of Botany, Faculty of Biology, University of Salamanca, E-37008 Salamanca (bernardos@usal. es) (2) Agricultural High School, Technical Institute of Bragan~a. P-5300 Bragan~a ([email protected]) (3) Herbarium/Botanic Garden, University ofTras-os-Montes e Alto Douro, P-5001-91 I Vila Real, ([email protected]) arrive le 10 juin 2004, accepte le 2 7 juillet 2004 Abstract.- The taxonomical composition and an analysis of four ecological cha racteristics of the 46 endemic species occurring in the lower Duero Basin (CW Iberian Peninsula) have been made. -

Ecological Assessments in the B+WISER Sites
Ecological Assessments in the B+WISER Sites (Northern Sierra Madre Natural Park, Upper Marikina-Kaliwa Forest Reserve, Bago River Watershed and Forest Reserve, Naujan Lake National Park and Subwatersheds, Mt. Kitanglad Range Natural Park and Mt. Apo Natural Park) Philippines Biodiversity & Watersheds Improved for Stronger Economy & Ecosystem Resilience (B+WISER) 23 March 2015 This publication was produced for review by the United States Agency for International Development. It was prepared by Chemonics International Inc. The Biodiversity and Watersheds Improved for Stronger Economy and Ecosystem Resilience Program is funded by the USAID, Contract No. AID-492-C-13-00002 and implemented by Chemonics International in association with: Fauna and Flora International (FFI) Haribon Foundation World Agroforestry Center (ICRAF) The author’s views expressed in this publication do not necessarily reflect the views of the United States Agency for International Development or the United States Government. Ecological Assessments in the B+WISER Sites Philippines Biodiversity and Watersheds Improved for Stronger Economy and Ecosystem Resilience (B+WISER) Program Implemented with: Department of Environment and Natural Resources Other National Government Agencies Local Government Units and Agencies Supported by: United States Agency for International Development Contract No.: AID-492-C-13-00002 Managed by: Chemonics International Inc. in partnership with Fauna and Flora International (FFI) Haribon Foundation World Agroforestry Center (ICRAF) 23 March -

Local and Landscape Compositions Influence Stingless Bee Communities and Pollination Networks in Tropical Mixed Fruit Orchards
diversity Article Local and Landscape Compositions Influence Stingless Bee Communities and Pollination Networks in Tropical Mixed Fruit Orchards, Thailand Kanuengnit Wayo 1 , Tuanjit Sritongchuay 2 , Bajaree Chuttong 3, Korrawat Attasopa 3 and Sara Bumrungsri 1,* 1 Department of Biology, Division of Biological Science, Faculty of Science, Prince of Songkla University, Hat Yai, Songkhla 90110, Thailand; [email protected] 2 Center for Integrative Conservation, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Menglun, Mengla 666303, China; [email protected] 3 Department of Entomology and Plant Pathology, Faculty of Agriculture, Chiang Mai University, Chiang Mai 50200, Thailand; [email protected] (B.C.); [email protected] (K.A.) * Correspondence: [email protected]; Tel.: +66-7428-8537 Received: 31 October 2020; Accepted: 15 December 2020; Published: 17 December 2020 Abstract: Stingless bees are vital pollinators for both wild and crop plants, yet their communities have been affected and altered by anthropogenic land-use change. Additionally, few studies have directly addressed the consequences of land-use change for meliponines, and knowledge on how their communities change across gradients in surrounding landscape cover remains scarce. Here, we examine both how local and landscape-level compositions as well as forest proximity affect both meliponine species richness and abundance together with pollination networks across 30 mixed fruit orchards in Southern Thailand. The results reveal that most landscape-level factors significantly influenced both stingless bee richness and abundance. Surrounding forest cover has a strong positive direct effect on both factors, while agricultural and urbanized cover generally reduced both bee abundance and diversity. In the local habitat, there is a significant interaction between orchard size and floral richness with stingless bee richness. -
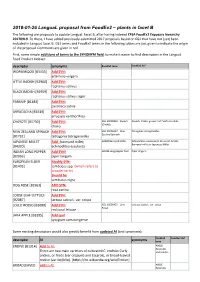
2018-01-26 Langual Proposal from Foodex2 – Plants in Facet B
2018-01-26 LanguaL proposal from FoodEx2 – plants in facet B The following are proposals to update LanguaL Facet B, after having indexed EFSA FoodEx2 Exposure hierarchy 20170919. To these, I have added previously-submitted 2017 proposals based on GS1 that have not (yet) been included in LanguaL facet B. GS1 terms and FoodEx2 terms in the following tables are just given to indicate the origin of the proposal. Comments are given in red. First, some simple additions of terms to the SYNONYM field, to make it easier to find descriptors in the LanguaL Food Product Indexer: descriptor synonyms FoodEx2 term FoodEx2 def WORMWOOD [B3433] Add SYN: artemisia vulgaris LITTLE RADISH [B2960] Add SYN: raphanus sativus BLACK RADISH [B2959] Add SYN: raphanus sativus niger PARSNIP [B1483] Add SYN: pastinaca sativa ARRACACHA [B3439] Add SYN: arracacia xanthorrhiza CHAYOTE [B1730] Add SYN: GS1 10006356 - Squash Squash, Choko, grown from Sechium edule (Choko) choko NEW ZEALAND SPINACH Add SYN: GS1 10006427 - New- Tetragonia tetragonoides Zealand Spinach [B1732] tetragonia tetragonoides JAPANESE MILLET Add : barnyard millet; A000Z Barnyard millet Echinochloa esculenta (A. Braun) H. Scholz, Barnyard millet or Japanese Millet. [B4320] echinochloa esculenta INDIAN LONG PEPPER Add SYN! A019B Long pepper fruit Piper longum [B2956] piper longum EUROPEAN ELDER Modify SYN: [B1403] sambucus spp. (which refers to broader term) Should be sambucus nigra DOG ROSE [B2961] ADD SYN: rosa canina LOOSE LEAF LETTUCE Add SYN: [B2087] lactusa sativa L. var. crispa LOLLO ROSSO [B2088] Add SYN: GS1 10006425 - Lollo Lactuca sativa L. var. crispa Rosso red coral lettuce JAVA APPLE [B3395] Add syn! syzygium samarangense Some existing descriptors would also greatly benefit from updated AI (and synonyms): FoodEx2 FoodEx2 def descriptor AI synonyms term ENDIVE [B1314] Add to AI: A00LD Escaroles There are two main varieties of cultivated C. -

Towards Preserving Threatened Grassland Species and Habitats
Towards preserving threatened grassland plant species and habitats - seed longevity, seed viability and phylogeography Dissertation zur Erlangung des Doktorgrades der Naturwissenschaften (Dr. rer. nat.) der Fakultät für Biologie und Vorklinische Medizin der Universität Regensburg vorgelegt von SIMONE B. TAUSCH aus Burghausen im Jahr 2017 II Das Promotionsgesuch wurde eingereicht am: 15.12.2017 Die Arbeit wurde angeleitet von: Prof. Dr. Peter Poschlod Regensburg, den 14.12.2017 Simone B. Tausch III IV Table of contents Chapter 1 General introduction 6 Chapter 2 Towards the origin of Central European grasslands: glacial and postgla- 12 cial history of the Salad Burnet (Sanguisorba minor Scop.) Chapter 3 A habitat-scale study of seed lifespan in artificial conditions 28 examining seed traits Chapter 4 Seed survival in the soil and at artificial storage: Implications for the 42 conservation of calcareous grassland species Chapter 5 How precise can X-ray predict the viability of wild flowering plant seeds? 56 Chapter 6 Seed dispersal in space and time - origin and conservation of calcareous 66 grasslands Summary 70 Zusammenfassung 72 References 74 Danksagung 89 DECLARATION OF MANUSCRIPTS Chapter 2 was published with the thesis’ author as main author: Tausch, S., Leipold, M., Poschlod, P. and Reisch, C. (2017). Molecular markers provide evidence for a broad-fronted recolonisation of the widespread calcareous grassland species Sanguisorba minor from southern and cryptic northern refugia. Plant Biology, 19: 562–570. doi:10.1111/plb.12570. V CHAPTER 1 General introduction THREATENED AND ENDANGERED persal ability (von Blanckenhagen & Poschlod, 2005). But in general, soils of calcareous grasslands exhibit HABITATS low ability to buffer species extinctions by serving as donor (Thompson et al., 1997; Bekker et al., 1998a; Regarding the situation of Europe’s plant species in- Kalamees & Zobel, 1998; Poschlod et al., 1998; Stöck- ventory, Central Europe represents the centre of en- lin & Fischer, 1999; Karlik & Poschlod, 2014). -

Syntaxonomical Revision of Quercetalia Pubescenti-Petraeae in the Italian Peninsula
Fitosociologia 41 (1): 87-164, 2004 87 Syntaxonomical revision of Quercetalia pubescenti-petraeae in the Italian Peninsula Blasi C.1, Di Pietro R.1 & Filesi L.2 1 Dipartimento di Biologia vegetale Università “La Sapienza” di Roma, P.le Aldo Moro 5, I-00185 Roma; e-mail: [email protected] 2 Dipartimento di Pianificazione, Università IUAV di Venezia, Cà Tron, Santa Croce 1957, I-30135 Venezia Abstract This paper presents a new sintaxonomical scheme for Quercetalia pubescenti-petraeae woodlands in peninsular Italy updated at the rank of sub- alliance. On the basis of bioclimatic, biogeographic and coenological diagnosis the Quercetalia pubescenti-petraeae order occurs in the study area in the form of five alliances: Quercion pubescenti-petraeae (suball.: Buxo-Quercenion pubescentis), Carpinion orientalis (suball.: Campanulo-Ostryenion, Laburno-Ostryenion stat. nov.; Lauro-Quercenion pubescentis and Cytiso-Quercenion pubescentis), Teucrio siculi-Quercion cerridis nom. conserv. propos. (suball.: Teucrio-Quercenion cerridis suball. nov., Ptilostemo-Quercenion cerridis); Pino-Quercion congestae (suball.: Pino-Quercenion congestae suball. nov., Quercenion virgilianae suball. nov.) and Erythronio-Quercion petraeae. The Quercion pubescenti-petraeae is a sintaxon with a sub-continental character and within the Italian Peninsula it is restricted to Ligurian-Piedmontese Apennine in the form of the sub-alliance Buxo- Quercenion pubescentis. Carpinion orientalis is a sintaxon with oceanic/suboceanic features, which is widely distributed along the entire Italian Peninsula. It occurs mainly on limestone substrates forming woodlands which can be potential vegetation types and also woods of secondary origin and evolution. The Teucrio-Quercion cerridis is an alliance which is widely distributed within central and southern Italy, where it finds its coenological optimum on acid, subacid or neutral substrates such as volcanic outcrops, flysch, sandstones or clayey-arenaceous sediments. -

Italy's Sibillini Mountains
Italy's Sibillini Mountains Naturetrek Tour Report 3 - 10 June 2015 Olive Skipper Lizard Orchid Report compiled by Philip Thompson Images courtesy of Stuart Morton Naturetrek Mingledown Barn Wolf's Lane Chawton Alton Hampshire GU34 3HJ UK T: +44 (0)1962 733051 F: +44 (0)1962 736426 E: [email protected] W: www.naturetrek.co.uk Italy's Sibillini Mountains Tour Report Tour Participants: Philip Thompson Tour Leader Sharon Thompson Co-leader With 13 Naturetrek clients Day 1 Wednesday 3rd June With our early flight safely completed, we met the early arrivals in the Arrivals Lounge of Ancona Airport and set off for the Sibillini National Park. We stopped en route at the Abbadia Di Fiastra Nature Reserve for an exploration and refreshments. After a coffee and soft drink, we undertook a walk around the perimeter of the park. Several bright green Italian Wall Lizards scuttled across the path and posed unconcernedly on the margins as we made our slow passage past. Numerous singing Blackcaps could be heard, joined by a couple of Nightingales deep within the woodland scrub. The pleasant purring sounds of a few Turtle Doves were joined by the melodious calls of a Golden Oriole, both of which eluded sighting. The temperature along the track was becoming oppressive and, joined with our slow progress, brought about the decision to retreat to the Monastery buildings and a light lunch! Whilst we ate we were able to enjoy the vibrant House Martin colony on the buildings with the Tree Sparrows and Common Redstarts on the rooftops and gutters. -

Federico Selvi a Critical Checklist of the Vascular Flora of Tuscan Maremma
Federico Selvi A critical checklist of the vascular flora of Tuscan Maremma (Grosseto province, Italy) Abstract Selvi, F.: A critical checklist of the vascular flora of Tuscan Maremma (Grosseto province, Italy). — Fl. Medit. 20: 47-139. 2010. — ISSN 1120-4052. The Tuscan Maremma is a historical region of central western Italy of remarkable ecological and landscape value, with a surface of about 4.420 km2 largely corresponding to the province of Grosseto. A critical inventory of the native and naturalized vascular plant species growing in this territory is here presented, based on over twenty years of author's collections and study of relevant herbarium materials and literature. The checklist includes 2.056 species and subspecies (excluding orchid hybrids), of which, however, 49 should be excluded, 67 need confirmation and 15 have most probably desappeared during the last century. Considering the 1.925 con- firmed taxa only, this area is home of about 25% of the Italian flora though representing only 1.5% of the national surface. The main phytogeographical features in terms of life-form distri- bution, chorological types, endemic species and taxa of particular conservation relevance are presented. Species not previously recorded from Tuscany are: Anthoxanthum ovatum Lag., Cardamine amporitana Sennen & Pau, Hieracium glaucinum Jord., H. maranzae (Murr & Zahn) Prain (H. neoplatyphyllum Gottschl.), H. murorum subsp. tenuiflorum (A.-T.) Schinz & R. Keller, H. vasconicum Martrin-Donos, Onobrychis arenaria (Kit.) DC., Typha domingensis (Pers.) Steud., Vicia loiseleurii (M. Bieb) Litv. and the exotic Oenothera speciosa Nutt. Key words: Flora, Phytogeography, Taxonomy, Tuscan Maremma. Introduction Inhabited by man since millennia and cradle of the Etruscan civilization, Maremma is a historical region of central-western Italy that stretches, in its broadest sense, from south- ern Tuscany to northern Latium in the provinces of Pisa, Livorno, Grosseto and Viterbo.