Manus-Pinus-Nigra.Pdf
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

BOE 015 De 18/01/2000 Sec 3 Pag 2271 a 2283
BOE núm. 15 Martes 18 enero 2000 2271 de Madrid, a través del Sistema Regional de Información y Promoción La Confederación Hidrográfica del Duero sometió el proyecto de la Tecnológica. Presa de Castrovido y trasvase del río Pedroso y el estudio de impacto Las tareas a desarrollar por la OTRI-ISCIII se concretan en: ambiental del mismo, conjuntamente, al trámite de información pública, mediante anuncio que se publicó en el «Boletín Oficial de la Provincia 1. Incorporación de la información de la Base de Datos a través de de Burgos» con fecha 2 de febrero de 1996, en virtud de lo establecido la aplicación informática suministrada por la Dirección General de Inves- en el artículo 15 del Reglamento. tigación. La Dirección General de Obras Hidráulicas y Calidad de las Aguas, 2. Difusión de toda la información del Sistema de Información Regio- en conformidad con el artículo 16 del Reglamento, remitió a la Dirección nal Madrid + D y facilitada por la Dirección General de Investigación. General de Calidad y Evaluación Ambiental, con fecha 26 de febrero de 3. Incorporación al sistema de información de investigadores que así 1997, el proyecto, estudio de impacto ambiental y el contenido de la infor- lo soliciten en el espacio de duración del Convenio. mación pública. El anexo II contiene los datos esenciales del proyecto. La información introducida o actualizada en el Sistema se utilizará Los aspectos más destacados del estudio de impacto ambiental se reco- para su explotación a través del Centro Virtual de Apoyo a la Innovación gen en el anexo III. Madrid + D. -

Listado De Vacantes
Página: 1/ 26 Listado de vacantes Cuerpo: 0597 MAESTROS Especialidad: AL AUDICIÓN Y LENGUAJE Loc. 090180002 ARANDA DE DUERO Centro: 09000148 CP INF-PRI CASTILLA Provincia: 09 Nº Vacante: 2 Tipo Vacante: ASIGNATURAS COMPLEMENTARIAS Asignaturas complementarias: 0597 EI EDUCACIÓN INFANTIL Loc. 090560001 BRIVIESCA Centro: 09000835 CP INF-PRI JUAN ABASCAL Provincia: 09 Nº Vacante: 56 Tipo Vacante: ITINERANTE + PROA Observaciones: ITINERA CON C.P "Dª MENCIA DE VELASCO" BRIVIESCA (BURGOS) Loc. 090590001 BURGOS Centro: 09000914 CP INF-PRI FRANCISCO DE VITORIA Provincia: 09 Nº Vacante: 95 Loc. 090590001 BURGOS Centro: 09001025 CP INF-PRI SOLAR DEL CID Provincia: 09 Nº Vacante: 233 Tipo Vacante: ITINERANTE + ASIGNATURAS COMPLEMENTARIAS Observaciones: ITINERA CON EL COLEGIO AURELIO GOMEZ ESCOLAR. Asignaturas complementarias: 0597 PT PEDAGOGÍA TERAPÉUTICA Loc. 091130001 COVARRUBIAS Centro: 09002388 CP INF-PRI ALEJANDRO RGUEZ. DE VALCARCEL Provincia: 09 Nº Vacante: 173 Tipo Vacante: ASIGNATURAS COMPLEMENTARIAS Observaciones: COMPARTIDA CON PRI. Asignaturas complementarias: 0597 PT PEDAGOGÍA TERAPÉUTICA Loc. 091510001 GUMIEL DE HIZAN Centro: 09002753 CP INF-PRI MAXIMINO S.MIGUEL DE LA CAMARA Provincia: 09 Nº Vacante: 201 Tipo Vacante: ASIGNATURAS COMPLEMENTARIAS Observaciones: COMPARTIDA CON PRI. Asignaturas complementarias: 0597 PT PEDAGOGÍA TERAPÉUTICA Loc. 099080012 QUINCOCES DE YUSO Centro: 09003290 CP INF-PRI VALLE DE LOSA Provincia: 09 Nº Vacante: 363 Tipo Vacante: ASIGNATURAS COMPLEMENTARIAS + PARCIAL Observaciones: COMPARTIDA CON PRI. Asignaturas complementarias: 0597 PT PEDAGOGÍA TERAPÉUTICA Loc. 092190011 MIRANDA DE EBRO Centro: 09003861 CP INF-PRI CERVANTES Provincia: 09 Nº Vacante: 320 Tipo Vacante: ITINERANTE Centros itinerantes: 09003897 CP INF-PRI LOS ÁNGELES (MIRANDA DE EBRO) Loc. 092890001 QUINTANAR DE LA SIERRA Centro: 09005018 CP INF-PRI VERA CRUZ Provincia: 09 Nº Vacante: 373 Tipo Vacante: ITINERANTE Centros itinerantes: 09005021 IES TIERRA DE ALVARGONZÁLEZ (QUINTANAR DE LA SIERRA) Loc. -

Provincia De BURGOS
— 47 — Provincia de BURGOS Comprende esta provincia los siguientes ayuntamientos por partidos judiciales: Partido de Aranda de Duero . Aguilera (La) . Fresnillo de las Dueñas . Pardilla. Tubilla del Lago . Aranda de Duero . Fuentelcésped . Peñalba de Castro . Vadocondes. Arandilla . Fuentenebro . Peñaranda de Duero. Valdeande. Baños de Valdearados . Fuentespina . Quemada. Vid (La) . Brazacorta. Gumiel de Hizán . Quintana del Pidio. Villalba de Duero. Caleruega. Gumiel del Mercado . San Juan del Monte . Villalbilla de Gumiel . Campillo de Aranda. Hontoria de Valdearados . Santa Cruz de la Salceda. Castrillo de la Vega . Milagros. Sotillo de la Ribera . Villanueva de Gumiel . Coruña del Conde . Oquillas. Corregalindo . Zazuar. Partido de Belorado. Alcocero. Espinosa del Camino. Pradoluengo . Valmala. Arraya de Oca. Eterna . Puras de Villafranca . Viloria de Rioja . Baseuñana Fresneda de la Sierra Tirón . Quintanaloranco. Villaescusa la Sombría. Belorado. Fresneda . Rábanos . Villafranca-Montes de Oca . Carrias. Castil de Carrias. Fresno de Riotirón . Redecilla del Camino. Villagalijo . Castildelgado . Garganchón . Redecilla del Campo . Villalbos. Cerezo de Riotirón. Ibrillos . San Clemente del Valle . Villalómez . Cerratón de Juarros . Ocón de Villafranca . Santa Cruz del Valle Urbión . Villambistia . Cueva-Cardiel . Pineda de la Sierra . Tosantos. Villanasur-Río de Oca. Partido de Briviesca. Abajas . Castil de Lences. Oña Rublacedo de abajo. Aguas Cándidas. Castil de Peones. Padrones de Bureba. Rucandio. Aguilar de Bureba . Cillaperlata. Parte de Bureba (La), Salas de Bureba . Bañuelos de Bureba. Cornudilla. Pino de Bureba. Salinillas de Bureba. Barcina de los Montes . Santa María del Invierno. Cubo de Bureba. Poza de la Sal. Barrios de Bureba (Los). Santa Olalla de Bureba . Frías. Prádanos de Bureba. Bentretea . Solas de Bureba . Quintanaélez . Berzosa de Bureba . -

Ruta De Las Setas
GASTRONOMÍA SERRANA La gastronomía serrana reúne, como no podía EL MÁGICO MUNDO DE ser menos, sabrosas recetas con setas y hongos, bien solos o como complementos de la carne. RUTA DE Tanto en primavera como en otoño, se celebran LAS SETAS Jornadas Micológicas en varias localidades de la Sierra de la Demanda. En ellas se realizan charlas La relación de setas que se dan —en abundancia— en esta ruta es amplia; La amanita cesárea, y coloquios, exposiciones, degustaciones y níscalo, champiñón silvestre, seta de cardo, de caballero, pie azul, boletus edulis, senderilla... muestras de estos variados frutos del bosque. LAS SETAS Las jornadas micológicas con mayor arraigo en la Los seteros saben bien que la Sierra de la Demanda es una zona óptima para practicar su afición. zona se celebran en Canicosa, Hontoria del Pinar Huellas de orígenes serranos, dinosaurios y La recolección de estos frutos del bosque es además, un complemento significativo en bastantes y Salas de los Infantes. necrópolis escondidos en bosques centenarios rentas familiares de la comarca. OTRAS ZONAS DE LA PROVINCIA También encontramos posibilidad de disfrutar de la recogida de setas y degustación en otras zonas de la provincia, como Villadiego, que ce- lebra anualmente jornadas micológicas al cobi- jo de los soportales de su Plaza Mayor o encon- trar diversos cotos en la zona de las Merindades. Jornadas micológicas FECHAS DENOMINACIÓN FERIA LOCALIDAD DIVERSIDAD DE HÁBITATS - VARIEDAD DE ESPECIES Mayo Jornadas Mico Ecológica / Tapa Mico Ecológica Comunero de Revenga La diversidad de hábitats (bosque, páramo, valle), se hace notar en esa variedad de especies micológicas. -

Bopbur-2014-084.Pdf
burgos boletín oficial de la provincia núm. 84 e martes, 6 de mayo de 2014 C.V.E.: BOPBUR-2014-084 sumario I. ADMINISTRACIÓN DEL ESTADO MINISTERIO DE AGRICULTURA, ALIMENTACIÓN Y MEDIO AMBIENTE CONFEDERACIÓN HIDROGRÁFICA DEL EBRO Comisaría de Aguas Solicitud de autorización de vertido de aguas residuales procedentes de la población de Casillas - Villarcayo de Merindad de Castilla la Vieja (Burgos) 5 II. ADMINISTRACIÓN AUTONÓMICA JUNTA DE CASTILLA Y LEÓN DELEGACIÓN TERRITORIAL DE BURGOS Servicio Territorial de Medio Ambiente Cambio de titularidad del coto de caza BU-10.811 de Fuentemolinos y Haza (Burgos) 6 Lotes de aprovechamientos de madera en montes de utilidad pública remitidos a las entidades propietarias, para su enajenación, en el primer trimestre de 2014 7 III. ADMINISTRACIÓN LOCAL DIPUTACIÓN PROVINCIAL DE BURGOS BIENESTAR SOCIAL Notificación de resolución sobre solicitudes de prestación económica de esta Diputación destinadas a la atención de personas con necesidades básicas de subsistencia en situaciones de urgencia social, ejercicio 2013 11 AYUNTAMIENTO DE ALCOCERO DE MOLA Concurso para el arrendamiento del aprovechamiento cinegético del coto de caza BU-10.792 12 diputación de burgos bopbur.diputaciondeburgos.es D.L.: BU -1-1958 –1– boletín oficial de la provincia burgos núm. 84 e martes, 6 de mayo de 2014 sumario AYUNTAMIENTO DE ARANDA DE DUERO SECRETARÍA GENERAL Aprobación definitiva de la ordenanza reguladora del control de acceso a las zonas peatonales 13 Corrección de errores en la convocatoria para la contratación de las -

NOMENCLÁTOR DE PARTIDOS JUDICIALES Sedes Y Municipios Integrantes
NOMENCLÁTOR DE PARTIDOS JUDICIALES Sedes y Municipios integrantes Cada Partido Judicial está formado por el conjunto de municipios que aparece agrupado con el mismo número. La cabecera, sede de los juzgados, figura en color rojo y el resto de municipios cuenta con un Juzgado de Paz. ANDALUCÍA PROVINCIA DE ALMERÍA Partido Judicial Código Municipio Población Nombre del Municipio 1 04001 1.248 ABLA 1 04002 1.183 ABRUCENA 1 04005 609 ALBOLODUY 1 04009 138 ALCUDIA DE MONTEAGUD 1 04010 677 ALHABIA 1 04011 3.691 ALHAMA DE ALMERÍA 1 04012 201 ALICÚN 1 04013 201.322 ALMERÍA 1 04014 176 ALMÓCITA 1 04015 125 ALSODUX 1 04023 120 BEIRES 1 04024 4.481 BENAHADUX 1 04026 58 BENITAGLA 1 04027 262 BENIZALÓN 1 04028 240 BENTARIQUE 1 04030 1.189 CANJÁYAR 1 04033 120 CASTRO DE FILABRES 1 04041 514 ENIX 1 04043 636 FELIX 1 04045 1.984 FIÑANA 1 04047 3.034 GÁDOR 1 04050 1.080 GERGAL 1 04051 486 HUÉCIJA 1 04052 17.917 HUÉRCAL DE ALMERÍA 1 04054 405 ILLAR 1 04055 448 INSTINCIÓN 1 04901 565 LAS TRES VILLAS 1 04060 564 LUCAINENA DE LAS TORRES 1 04065 496 NACIMIENTO 1 04066 31.666 NÍJAR 1 04067 558 OHANES 1 04068 191 OLULA DE CASTRO 1 04071 414 PADULES 1 04074 4.103 PECHINA 1 04077 303 RÁGOL 1 04078 1.453 RIOJA 1 04080 199 SANTA CRUZ DE MARCHENA 1 04081 461 SANTA FE DE MONDÚJAR 1 04082 289 SENÉS 1 04086 2.436 SORBAS 1 04088 3.717 TABERNAS Página 1 12/02/2021 1 04090 336 TAHAL 1 04091 365 TERQUE 1 04094 248 TURRILLAS 1 04095 822 ULEILA DEL CAMPO 1 04097 230 VELEFIQUE 1 04101 5.978 VIATOR 1 04102 26.899 VÍCAR Total Población Partido Judicial: 324.637 2 04003 25.412 -

Portada Acta 29
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Diposit Digital de la Universitat de Barcelona J. I. PADILLA* K. A. RUEDA** EL DESPOBLADO ALTOMEDIEVAL DE CUYACABRAS (BURGOS): REALIDAD, PRINCIPIOS Y ARGUMENTOS ABSTRACT The archaeological site of Cuyacabras (Quintanar de la Sierra, Burgos) is one of main early medieval reference settlements in Upper Arlanza basin lands and it attracts a great amount of visitors. The monumental featured half-caved church and the necropolis consisting of several burials dug into the rock present around the whole area are material evidences of an ancient hill town which has remained uninhabited from middle XIIIth century. New fi eldworks on this whole site, excavated during the sixties, have been fo- cused on improving the scientifi c impact of the site and setting up to date the records and information entries for the necropolis. All obtained data emphasize the weakness of several principles used to put in value the settlement, and they also suggest a revision of several-year-old discussions on one hand, and on the other one, they encourage scholars to propound new up to date theories which will fi t better with our knowledge’s current context. El lugar de Cuyacabras se encuentra situado en un paraje forestal perteneciente al término municipal de Quintanar de la Sierra, en pleno corazón de la comarca natural de Pinares y sobre las estribaciones meridionales de la sierra de la Demanda. Este yacimiento arqueológico, que parece esconderse entre los pinares de este mu- nicipio burgalés, es uno de los conjuntos arqueológicos más conocidos y visitados *Professor Titular d’Historia Medieval. -

Situacion De La Concentración Parcelaria En La Provincia De Burgos
SITUACION DE LA CONCENTRACIÓN PARCELARIA EN LA PROVINCIA DE BURGOS 45 ANALISIS DE LA SITUACION DE LA CONCENTRACIÓN PARCELARIA EN LA PROVINCIA DE BURGOS 1.1 SITUACIÓN DE LA CONCENTRACIÓN PARCELARIA: La situación de la concentración parcelaria en la provincia de Burgos, en porcentaje respecto a la superficie concentrable, y sin tener en cuenta reconcentraciones, se puede comprobar en el siguiente gráfico: SITUACIÓN DE LA CONCENTRACIÓN PARCELARIA DE LA PROVINCIA DE BURGOS EN % RESPECTO A LA SUPERFICIE CONCENTRABLE SUPERFICIE SOLICITADA EN TRABAJOS PREVIOS AL ACUERDO DE SUPERFICIE UTILIDAD PÚBLICA FINALIZADA+ACTUANDO 2,5% 73,7% SUPERFICIE ACTUANDO 5,5% SUPERFICIE SOLICITADA NO ACTUANDO 2,6% SUPERFICIE FINALIZADA SUPERFICIE NO 68,2% SOLICITADA 21,2% En la provincia de Burgos, la superficie concentrada (68,2%) más la superficie que se está concentrando (5,5%) supone un 73,7% de la superficie concentrable de la provincia. La superficie solicitada asciende al 5,1%, estando un 2,5% de la superficie en trabajos previos al acuerdo que declara la utilidad pública de la concentración parcelaria: tramitación de Estudio Técnico Previo o tramitación ambiental (evaluación de impacto ambiental o resolución sobre sometimiento/no sometimiento a evaluación de impacto ambiental). En el resto de zonas solicitadas, que representan un 2,6% de la superficie concentrable, no se ha iniciado ninguna actuación en aplicación de las prioridades establecidas o en algunos casos se ha constatado un ambiente desfavorable al inicio de actuaciones. La superficie no solicitada -

Macrorremains Evidence of Anthropogenic Recession of Pinus
Macrorremains evidence of anthropogenic recession of Pinus nigra Aiton in Northern Spain García-Amorena Ignacio, Moreno Amat, E, Iglesias González R, Rubiales JM, Gómez-Manzaneque F E.T.S.I. Montes (Dpto. Silvopascicultura), Universidad Politécnica de Madrid. Avda. Ramiro de Maeztu s/n, 28040 Madrid CONTACT: [email protected] Introduction Study sites The natural forest landscape of the Duero Basin in Spain’s Northern Meseta is now reduced to small masses, its demise largely owed to the intense STUDY SITES (rombs) agricultural and stock-raising use made of its lands. The A-Tubilla del Agua composition of the scant natural masses suggests that B-Tubilla del Lago, formations dominated by broadleaved species once C-Fuentetoba occupied a much greater area. This idea has led to the believe that pine trees have few 1: Espinosa de Cerrato (Franco Múgica et al., 2001) 2: La Piedra (Ramil-Rego et al., 1998) place within the natural dynamics of this area (Carrión 3: Valle de la Nava (Menéndez Amor, 1968) and Fernández, 2009). However, P. pinaster Aiton and P. 4: Quinatanar de la Sierra (Peñalba et al., 1997) pinea L. on low-land sandy areas, P. sylvestris L. and P. 5: el Hornillo (Gómez-Lobo, 1993) uncinata Ramond ex DC on mountain high areas have 6: Quintana Redonda (García Antón et al., 1995). 7: Aguilar de Campoo (Alcalde et al., 2001) been demonstrated to have played an important role 8: Cevico Navero (Roig et al., 1997) during the Holocene (Franco et al., 2005; Rubiales et al., 2010). Orange: The natural distribution of P. -
Cadenza Document
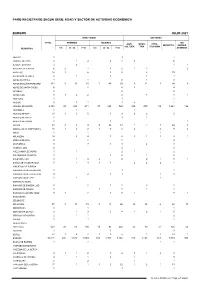
PARO REGISTRADO SEGÚN SEXO, EDAD Y SECTOR DE ACTIVIDAD ECONÓMICA BURGOS JULIO 2021 SEXO Y EDAD SECTORES TOTAL HOMBRES MUJERES SIN AGRI- INDUS- CONS- SERVICIOS EMPLEO CULTURA TRIA TRUCCIÓN MUNICIPIOS <25 25 - 44 >=45 <25 25 - 44 >=45 ANTERIOR ABAJAS 1 1 1 ADRADA DE HAZA 9 1 2 2 4 3 6 AGUAS CANDIDAS 2 2 1 1 AGUILAR DE BUREBA 6 2 1 1 2 1 3 2 ALBILLOS 12 1 6 1 4 2 10 ALCOCERO DE MOLA 2 1 1 1 1 ALFOZ DE BRICIA 3 1 2 1 2 ALFOZ DE QUINTANADUEÑA 122 5 13 20 5 40 39 4 11 5 94 8 ALFOZ DE SANTA GADEA 6 2 4 2 4 ALTABLE 2 2 2 ALTOS, LOS 7 1 2 2 2 3 3 1 AMEYUGO 1 1 1 ANGUIX 5 3 1 1 4 1 ARANDA DE DUERO 2.151 98 288 411 87 538 729 339 270 89 1.261 192 ARANDILLA 1 1 1 ARAUZO DE MIEL 13 1 3 5 1 3 6 2 5 ARAUZO DE SALCE 1 1 1 ARAUZO DE TORRE 1 1 1 ARCOS 58 4 5 9 4 24 12 1 7 3 44 3 ARENILLAS DE RIOPISUERGA 16 1 2 2 1 6 4 4 2 8 2 ARIJA 1 1 1 ARLANZON 14 3 4 3 4 2 1 2 8 1 ARRAYA DE OCA 4 2 2 4 ATAPUERCA 5 3 1 1 2 3 AUSINES, LOS 2 2 2 AVELLANOSA DE MUÑO 3 1 2 1 2 BAHABON DE ESGUEVA 3 1 2 1 2 BALBASES, LOS 9 1 4 2 2 2 1 5 1 BAÑOS DE VALDEARADOS 7 1 4 1 1 2 2 2 1 BAÑUELOS DE BUREBA BARBADILLO DE HERREROS 1 1 1 BARBADILLO DEL MERCADO 4 2 1 1 1 3 BARBADILLO DEL PEZ 1 1 1 BARRIO DE MUÑO BARRIOS DE BUREBA, LOS 5 2 1 2 3 2 BARRIOS DE COLINA 3 1 2 1 2 BASCONCILLOS DEL TOZO 9 1 2 1 5 1 7 1 BASCUÑANA BELBIMBRE BELORADO 87 8 13 19 3 18 26 12 14 6 42 13 BERBERANA 1 1 1 BERLANGAS DE ROA 9 1 3 1 4 1 2 1 5 BERZOSA DE BUREBA 2 1 1 2 BOZOO 1 1 1 BRAZACORTA BRIVIESCA 521 25 65 100 14 93 224 25 89 37 325 45 BUGEDO 3 3 3 BUNIEL 27 1 4 7 1 9 5 1 2 23 1 BURGOS 10.018 482 1.510 2.069 -

Maquetación 1
boletín oficial de la provincia burgos núm. 221 e miércoles, 20 de noviembre de 2013 C.V.E.: BOPBUR-2013-08513 III. ADMINISTRACIÓN LOCAL DIPUTACIÓN PROVINCIAL DE BURGOS RECAUDACIÓN DE TRIBUTOS Citación para ser notificados por comparecencia Procedimiento. – Recibos pendientes del ejercicio 2012 del siguiente concepto: Liquidación de ingreso directo de: IBIUL Impuesto bienes inmuebles. Urbana (liquidaciones). TTRESI ID Tratamiento de residuos sólidos (liquidaciones). IIVT Impuesto incremento valor de los terrenos. La Diputación Provincial, en ejercicio de sus competencias, y no habiendo resultado posible practicar la notificación al interesado o a su representante, por causas no impu- tables a esta Administración, intentada en cumplimiento de lo dispuesto en el artículo 112 de la Ley General Tributaria 58/2003, de 17 de diciembre, mediante el presente anuncio se cita a los contribuyentes abajo indicados para que comparezcan en el lugar que asimismo se indica, a fin de que les sean notificadas las actuaciones llevadas a cabo en los proce- dimientos tributarios que les afectan. Lugar y plazo de comparecencia. – Las personas interesadas, o sus representantes, deberán comparecer, con el fin de ser notificadas, en las oficinas de este órgano administrativo, ubicadas en el Paseo del Espolón, n.º 34 (Palacio Provincial) en el plazo de quince (15) días, a contar desde el siguiente al de la publicación de este anuncio en el «Boletín Oficial» de la provincia. Se advierte a los interesados que, de no comparecer en dicho plazo, la notificación se entenderá practicada, a todos los efectos legales, desde el día siguiente al del vencimiento del mismo. Burgos, a 6 de noviembre de 2013. -

Lo Que Fue Y Es Quintanar Para Quintanar De La Sierra
TOMAS MEDRANO LAZARO LO QUE FUE Y ES QUINTANAR PARA QUINTANAR DE LA SIERRA ANO DE 1950 J TÜ1WAS IttEDKAMÜ lAZARII LO QUE FUE Y ES QUINTAM Y PARA OUINTAMR UE LA SIERRA A iÑI O'AD E 19 50 PROLOGO Confieso que tanto en esta ocasión, como en otra hace cerca de cuarenta años en que escribí un sencillo libro sobre mi patria chica, cuyo titulo era A mi desgraciado pueblo, mis sudores me ha costado, además de demostrar que yo no he nacido para profeta, porque desde aquella fecha en que parece veia yo a mi querido pueblo amenazado de una ruina inevitable, aquel pesimismo que yo forjaba ha sido el re• verso de la medalla, y nadie puede suponer lo que celebro mi equivocación, ya que la riqueza de los pueblos se mide por su número de habitantes y esta nuestra querida villa se ha puesto en ese lapso de tiempo como las primeras de la provincia. Cuánto gozaría yo, si otras personas más cultas y desde luego más autorizadas, escribieran algo de mi pueblo, de su naturaleza, de manera que quedara ya formado en pequeño el principio de una futura, útil y sencilla Biblioteca. ¡Con qué satisfacción, con qué ansia devoraría yo esa obrita! Pero nada, está visto que esas personas capacitadas para esta clase de trabajos rehuyen estimular nuestros progre• sos, estudiar nuestras dotes o nuestras miserias. ¡Cuánto lo celebraría Quintanar, que bien se lo merece! El móvil que — 4 — me guía a escribir este librito, como decía en mi anterior, no es otro que el excesivo cariño que tengo a mi querido pueblo, pidiendo benevolencia a todos mis lectores y rogán• doles dispensen mi osadía, pues aun cuando no encuentren ningún mérito, porque no le tiene, vean la mucha volun• tad en un bijo del pueblo, que no vió más letras que las que aprendió en la Escuela del mismo.