Moringa Oleifera Extract Extenuates Echis Ocellatus Venom-Induced Toxicities, Histopathological Impairments and Inflammation Via Enhancement of Nrf2 Expression in Rats
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Snake Bite Protocol
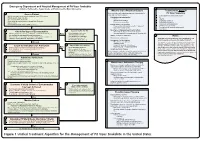
Lavonas et al. BMC Emergency Medicine 2011, 11:2 Page 4 of 15 http://www.biomedcentral.com/1471-227X/11/2 and other Rocky Mountain Poison and Drug Center treatment of patients bitten by coral snakes (family Ela- staff. The antivenom manufacturer provided funding pidae), nor by snakes that are not indigenous to the US. support. Sponsor representatives were not present dur- At the time this algorithm was developed, the only ing the webinar or panel discussions. Sponsor represen- antivenom commercially available for the treatment of tatives reviewed the final manuscript before publication pit viper envenomation in the US is Crotalidae Polyva- ® for the sole purpose of identifying proprietary informa- lent Immune Fab (ovine) (CroFab , Protherics, Nash- tion. No modifications of the manuscript were requested ville, TN). All treatment recommendations and dosing by the manufacturer. apply to this antivenom. This algorithm does not con- sider treatment with whole IgG antivenom (Antivenin Results (Crotalidae) Polyvalent, equine origin (Wyeth-Ayerst, Final unified treatment algorithm Marietta, Pennsylvania, USA)), because production of The unified treatment algorithm is shown in Figure 1. that antivenom has been discontinued and all extant The final version was endorsed unanimously. Specific lots have expired. This antivenom also does not consider considerations endorsed by the panelists are as follows: treatment with other antivenom products under devel- opment. Because the panel members are all hospital- Role of the unified treatment algorithm -

Medically Important Differences in Snake Venom Composition Are Dictated by Distinct Postgenomic Mechanisms
Medically important differences in snake venom composition are dictated by distinct postgenomic mechanisms Nicholas R. Casewella,b,1, Simon C. Wagstaffc, Wolfgang Wüsterb, Darren A. N. Cooka, Fiona M. S. Boltona, Sarah I. Kinga, Davinia Plad, Libia Sanzd, Juan J. Calveted, and Robert A. Harrisona aAlistair Reid Venom Research Unit and cBioinformatics Unit, Liverpool School of Tropical Medicine, Liverpool L3 5QA, United Kingdom; bMolecular Ecology and Evolution Group, School of Biological Sciences, Bangor University, Bangor LL57 2UW, United Kingdom; and dInstituto de Biomedicina de Valencia, Consejo Superior de Investigaciones Científicas, 11 46010 Valencia, Spain Edited by David B. Wake, University of California, Berkeley, CA, and approved May 14, 2014 (received for review March 27, 2014) Variation in venom composition is a ubiquitous phenomenon in few (approximately 5–10) multilocus gene families, with each snakes and occurs both interspecifically and intraspecifically. family capable of producing related isoforms generated by Venom variation can have severe outcomes for snakebite victims gene duplication events occurring over evolutionary time (1, 14, by rendering the specific antibodies found in antivenoms in- 15). The birth and death model of gene evolution (16) is fre- effective against heterologous toxins found in different venoms. quently invoked as the mechanism giving rise to venom gene The rapid evolutionary expansion of different toxin-encoding paralogs, with evidence that natural selection acting on surface gene families in different snake lineages is widely perceived as the exposed residues of the resulting gene duplicates facilitates main cause of venom variation. However, this view is simplistic subfunctionalization/neofunctionalization of the encoded proteins and disregards the understudied influence that processes acting (15, 17–19). -

High Throughput Screening and Identification of Coagulopathic Snake Venom Proteins and 2 Peptides Using Nanofractionation and Proteomics Approaches
bioRxiv preprint doi: https://doi.org/10.1101/780155; this version posted September 23, 2019. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY 4.0 International license. Classified Personnel Information 1 High throughput screening and identification of coagulopathic snake venom proteins and 2 peptides using nanofractionation and proteomics approaches 3 4 Julien Slagbooma,b, Marija Mladićc, Chunfang Xie b, Freek Vonkd, Govert W. Somsenb, Nicholas 5 R. Casewella, Jeroen Koolb 6 aCentre for Snakebite Research & Interventions, Liverpool School of Tropical Medicine, 7 Liverpool, UK 8 bDivision of BioAnalytical Chemistry, Amsterdam Institute for Molecules Medicines and Systems, 9 VU University Amsterdam, Amsterdam, The Netherlands 10 cAnimal Sciences and Health, Institute of Biology Leiden, Leiden University, Leiden, The 11 Netherlands 12 dNaturalis Biodiversity Center, Leiden, The Netherlands 13 *Corresponding author [email protected] 14 15 1 bioRxiv preprint doi: https://doi.org/10.1101/780155; this version posted September 23, 2019. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY 4.0 International license. Classified Personnel Information 16 Abstract 17 Snakebite is a neglected tropical disease that results in a variety of systemic and local pathologies in 18 envenomed victims and is responsible for around 138,000 deaths every year. Many snake venoms cause 19 severe coagulopathy that makes victims vulnerable to suffering life-threating haemorrhage. -

Long-Term Effects of Snake Envenoming
toxins Review Long-Term Effects of Snake Envenoming Subodha Waiddyanatha 1,2, Anjana Silva 1,2 , Sisira Siribaddana 1 and Geoffrey K. Isbister 2,3,* 1 Faculty of Medicine and Allied Sciences, Rajarata University of Sri Lanka, Saliyapura 50008, Sri Lanka; [email protected] (S.W.); [email protected] (A.S.); [email protected] (S.S.) 2 South Asian Clinical Toxicology Research Collaboration, Faculty of Medicine, University of Peradeniya, Peradeniya 20400, Sri Lanka 3 Clinical Toxicology Research Group, University of Newcastle, Callaghan, NSW 2308, Australia * Correspondence: [email protected] or [email protected]; Tel.: +612-4921-1211 Received: 14 March 2019; Accepted: 29 March 2019; Published: 31 March 2019 Abstract: Long-term effects of envenoming compromise the quality of life of the survivors of snakebite. We searched MEDLINE (from 1946) and EMBASE (from 1947) until October 2018 for clinical literature on the long-term effects of snake envenoming using different combinations of search terms. We classified conditions that last or appear more than six weeks following envenoming as long term or delayed effects of envenoming. Of 257 records identified, 51 articles describe the long-term effects of snake envenoming and were reviewed. Disability due to amputations, deformities, contracture formation, and chronic ulceration, rarely with malignant change, have resulted from local necrosis due to bites mainly from African and Asian cobras, and Central and South American Pit-vipers. Progression of acute kidney injury into chronic renal failure in Russell’s viper bites has been reported in several studies from India and Sri Lanka. Neuromuscular toxicity does not appear to result in long-term effects. -

Snakebite: the World's Biggest Hidden Health Crisis
Snakebite: The world's biggest hidden health crisis Snakebite is a potentially life-threatening neglected tropical disease (NTD) that is responsible for immense suffering among some 5.8 billion people who are at risk of encountering a venomous snake. The human cost of snakebite Snakebite Treatment Timeline Each year, approximately 5.4 million people are bitten by a snake, of whom 2.7 million are injected with venom. The first snake antivenom This leads to 400,000 people being permanently dis- produced, against the Indian Cobra. abled and between 83,000-138,000 deaths annually, Immunotherapy with animal- mostly in sub-Saharan Africa and South Asia. 1895 derived antivenom has continued to be the main treatment for snakebite evenoming for 120 years Snakebite: both a consequence and a cause of tropical poverty The Fav-Afrique antivenom, 2014 produced by Sanofi Pasteur (France) Survivors of untreated envenoming may be left with permanently discontinued amputation, blindness, mental health issues, and other forms of disability that severely affect their productivity. World Health Organization Most victims are agricultural workers and children in 2018 (WHO) lists snakebite envenoming the poorest parts of Africa and Asia. The economic as a neglected tropical disease cost of treating snakebite envenoming is unimaginable in most communities and puts families and communi- ties at risk of economic peril just to pay for treatment. WHO launches a strategy to prevent and control snakebite envenoming, including a program targeting affected communities and their health systems Global antivenom crisis 2019 The world produces less than half of the antivenom it The Scientific Research Partnership needs, and this only covers 57% of the world’s species for Neglected Tropical Snakbites of venomous snake. -

Bitis Arietans) Venom, and Their Neutralization by Antivenom
Toxins 2014, 6, 1586-1597; doi:10.3390/toxins6051586 OPEN ACCESS toxins ISSN 2072-6651 www.mdpi.com/journal/toxins Article In Vitro Toxic Effects of Puff Adder (Bitis arietans) Venom, and Their Neutralization by Antivenom Steven Fernandez 1, Wayne Hodgson 1, Janeyuth Chaisakul 2, Rachelle Kornhauser 1, Nicki Konstantakopoulos 1, Alexander Ian Smith 3 and Sanjaya Kuruppu 3,* 1 Department of Pharmacology, Monash University, Building 13E, Wellington Road, Clayton, Vic 3800, Australia; E-Mails: [email protected] (S.F.); [email protected] (W.H.); [email protected] (R.K.); [email protected] (N.K.) 2 Department of Pharmacology, Phramongkutklao College of Medicine, Bangkok 10400, Thailand; E-Mail: [email protected] 3 Department of Biochemistry & Molecular Biology, Monash University, Building 77, Wellington Road, Clayton, Vic 3800, Australia; E-Mail: [email protected] * Author to whom correspondence should be addressed; E-Mail: [email protected]; Tel: +61-3-9902-9372; Fax: + 61-3-9902-9500. Received: 24 November 2013; in revised form: 6 April 2014 / Accepted: 4 May 2014 / Published: 19 May 2014 Abstract: This study investigated the in vitro toxic effects of Bitis arietans venom and the ability of antivenom produced by the South African Institute of Medical Research (SAIMR) to neutralize these effects. The venom (50 µg/mL) reduced nerve-mediated twitches of the chick biventer muscle to 19% ± 2% of initial magnitude (n = 4) within 2 h. This inhibitory effect of the venom was significantly attenuated by prior incubation of tissues with SAIMR antivenom (0.864 µg/µL; 67% ± 4%; P < 0.05; n = 3–5, unpaired t-test). -

Testing the Toxicofera: Comparative Reptile Transcriptomics Casts Doubt on the Single, Early Evolution of the Reptile Venom Syst
bioRxiv preprint doi: https://doi.org/10.1101/006031; this version posted June 6, 2014. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. 1 Title page 2 3 Title: 4 Testing the Toxicofera: comparative reptile transcriptomics casts doubt on the single, early 5 evolution of the reptile venom system 6 7 Authors: 8 Adam D Hargreaves1, Martin T Swain2, Darren W Logan3 and John F Mulley1* 9 10 Affiliations: 11 1. School of Biological Sciences, Bangor University, Brambell Building, Deiniol Road, 12 Bangor, Gwynedd, LL57 2UW, United Kingdom 13 14 2. Institute of Biological, Environmental & Rural Sciences, Aberystwyth University, 15 Penglais, Aberystwyth, Ceredigion, SY23 3DA, United Kingdom 16 17 3. Wellcome Trust Sanger Institute, Hinxton, Cambridge, CB10 1HH, United Kingdom 18 19 *To whom correspondence should be addressed: [email protected] 20 21 22 Author email addresses: 23 John Mulley - [email protected] 24 Adam Hargreaves - [email protected] 25 Darren Logan - [email protected] 26 Martin Swain - [email protected] 1 bioRxiv preprint doi: https://doi.org/10.1101/006031; this version posted June 6, 2014. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. -

Medicines/Pharmaceuticals of Animal Origin V3.0 November 2020
Medicines/pharmaceuticals of animal origin V3.0 November 2020 Medicines/pharmaceuticals of animal origin - This guideline provides information for all clinical staff within Hospital and Health Services (HHS) on best practice for avoidance of issues related to animal products. Medicines/pharmaceuticals of animal origin - V3.0 November 2020 Published by the State of Queensland (Queensland Health), November 2020 This document is licensed under a Creative Commons Attribution 3.0 Australia licence. To view a copy of this licence, visit creativecommons.org/licenses/by/3.0/au © State of Queensland (Queensland Health) 2020 You are free to copy, communicate and adapt the work, as long as you attribute the State of Queensland (Queensland Health). For more information contact: Medication Services Queensland, Queensland Health, GPO Box 48, Brisbane QLD 4001, email [email protected] An electronic version of this document is available at https://www.health.qld.gov.au/__data/assets/pdf_file/0024/147507/qh-gdl-954.pdf Disclaimer: The content presented in this publication is distributed by the Queensland Government as an information source only. The State of Queensland makes no statements, representations or warranties about the accuracy, completeness or reliability of any information contained in this publication. The State of Queensland disclaims all responsibility and all liability (including without limitation for liability in negligence) for all expenses, losses, damages and costs you might incur as a result of the information being inaccurate -

Epidemiology and Clinical Outcomes of Snakebite in the Elderly: a Toxic Database Study
Clinical Toxicology ISSN: 1556-3650 (Print) 1556-9519 (Online) Journal homepage: http://www.tandfonline.com/loi/ictx20 Epidemiology and clinical outcomes of snakebite in the elderly: a ToxIC database study Meghan B. Spyres, Anne-Michelle Ruha, Kurt Kleinschmidt, Rais Vohra, Eric Smith & Angela Padilla-Jones To cite this article: Meghan B. Spyres, Anne-Michelle Ruha, Kurt Kleinschmidt, Rais Vohra, Eric Smith & Angela Padilla-Jones (2018) Epidemiology and clinical outcomes of snakebite in the elderly: a ToxIC database study, Clinical Toxicology, 56:2, 108-112, DOI: 10.1080/15563650.2017.1342829 To link to this article: https://doi.org/10.1080/15563650.2017.1342829 Published online: 13 Jul 2017. Submit your article to this journal Article views: 120 View related articles View Crossmark data Full Terms & Conditions of access and use can be found at http://www.tandfonline.com/action/journalInformation?journalCode=ictx20 CLINICAL TOXICOLOGY, 2018 VOL. 56, NO. 2, 108–112 https://doi.org/10.1080/15563650.2017.1342829 CLINICAL RESEARCH Epidemiology and clinical outcomes of snakebite in the elderly: a ToxIC Ã database study Meghan B. Spyresa, Anne-Michelle Ruhab, Kurt Kleinschmidtc, Rais Vohrad, Eric Smithc and Angela Padilla-Jonesb aDepartment of Emergency Medicine, Division of Medical Toxicology, University of Southern California, Los Angeles, CA, USA; bDepartment of Medical Toxicology, Banner-University Medical Center Phoenix, Phoenix, AZ, USA; cDepartment of Emergency Medicine, Univesity of Texas Southwestern Medical Center, Dallas, TX, USA; dDepartment of Emergency Medicine, UCSF Fresno Medical Center, Fresno, CA, USA ABSTRACT ARTICLE HISTORY Introduction: Epidemiologic studies of snakebites in the United States report typical victims to be Received 23 February 2017 young men. -

Polyvalent Horse F(Ab`)2 Snake Antivenom: Development of Process to Produce Polyvalent Horse F(Ab`)2 Antibodies Anti-African Snake Venom
African Journal of Biotechnology Vol. 9 (16), pp. 2446-2455, 19 April, 2010 Available online at http://www.academicjournals.org/AJB ISSN 1684–5315 © 2009 Academic Journals Full Length Research Paper Polyvalent horse F(Ab`)2 snake antivenom: Development of process to produce polyvalent horse F(Ab`)2 antibodies anti-african snake venom R. G. Guidlolin1, R. M. Marcelino1, H. H. Gondo1, J. F. Morais1, R. A. Ferreira2, C. L. Silva2, T. L. Kipnis2, J. A. Silva2, J. Fafetine3 and W. D. da Silva2* 1Divisao Bioindustrial, Instituto Butantan, Sao Paulo, SP, Brazil. 2Laboratório de Biologia do Reconhecer, Centro de Biociencias e Biotecnologia, Universidade Estadual do Norte Fluminense – Darcy Ribeiro, Campos dos Goytacazes, RJ, Brazil. 3Universidade Eduardo Mondlane, Maputo, Mozambique. Accepted 23 March, 2010 A method to obtain polyvalent anti-Bitis and polyvalent-anti-Naja antibodies was developed by immunizing horses with B. arietans, B. nasicornis, B. rhinoceros, N. melanoleuca and N. mossambica crude venoms. Antibody production was followed by the ELISA method during the immunization procedure. Once the desired anti-venom antibody titers were attained, horses were bled and the immunoglobulins were separated from the sera by (NH4)2SO4 precipitation, cleaved with pepsin and filtered through a 30 kDa ultrafiltration membrane. F(ab´)2 fragments were further purified by Q-Fast Flow chromatography, concentrated by molecular ultrafiltration and sterilized by filtration through 0.22 m membranes. The resulting F(ab´)2 preparations were rich in intact L and in pieces of H IgG(T) chains, as demonstrated by electrophoresis and Western blot and exhibited high antibody titers, as assayed by the ELISA method. -

The Snakes of Niger
Official journal website: Amphibian & Reptile Conservation amphibian-reptile-conservation.org 9(2) [Special Section]: 39–55 (e110). The snakes of Niger 1Jean-François Trape and Youssouph Mané 1Institut de Recherche pour le Développement (IRD), UMR MIVEGEC, Laboratoire de Paludologie et de Zoologie Médicale, B.P. 1386, Dakar, SENEGAL Abstract.—We present here the results of a study of 1,714 snakes from the Republic of Niger, West Africa, collected from 2004 to 2008 at 28 localities within the country. Based on this data, supplemented with additional museum specimens (23 selected specimens belonging to 10 species) and reliable literature reports, we present an annotated checklist of the 51 snake species known from Niger. Psammophis sudanensis is added to the snake fauna of Niger. Known localities for all species are presented and, where necessary, taxonomic and biogeographic issues discussed. Key words. Reptilia; Squamata; Ophidia; taxonomy; biogeography; species richness; venomous snakes; Niger Re- public; West Africa Citation: Trape J-F and Mané Y. 2015. The snakes of Niger. Amphibian & Reptile Conservation 9(2) [Special Section]: 39–55 (e110). Copyright: © 2015 Trape and Mané. This is an open-access article distributed under the terms of the Creative Commons Attribution-NonCommercial- NoDerivatives 4.0 International License, which permits unrestricted use for non-commercial and education purposes only, in any medium, provided the original author and the official and authorized publication sources are recognized and properly credited. -

Cost-Effectiveness of Antivenoms for Snakebite Envenoming in 16 Countries in West Africa
RESEARCH ARTICLE Cost-Effectiveness of Antivenoms for Snakebite Envenoming in 16 Countries in West Africa Muhammad Hamza1, Maryam A. Idris1, Musa B. Maiyaki1, Mohammed Lamorde2, Jean- Philippe Chippaux3, David A. Warrell4, Andreas Kuznik1,2,5, Abdulrazaq G. Habib1* 1 College of Health of Sciences, Bayero University, Kano, Nigeria, 2 Infectious Diseases Institute, Makerere University College of Health Sciences, Kampala, Uganda, 3 Institut de Recherche pour le Development, Cotonou, Benin Republic and Université Paris Descartes, Sorbonne Paris Cité, Faculté de Pharmacie, Paris, France, 4 Nuffield Department of Medicine, University of Oxford, Oxford, United Kingdom, 5 Celgene Corporation, Warren, New Jersey, United States of America * [email protected] Abstract Background Snakebite poisoning is a significant medical problem in agricultural societies in Sub Saha- OPEN ACCESS ran Africa. Antivenom (AV) is the standard treatment, and we assessed the cost-effective- Citation: Hamza M, Idris MA, Maiyaki MB, Lamorde ness of making it available in 16 countries in West Africa. M, Chippaux J-P, Warrell DA, et al. (2016) Cost- Effectiveness of Antivenoms for Snakebite Envenoming in 16 Countries in West Africa. PLoS Methods Negl Trop Dis 10(3): e0004568. doi:10.1371/journal. We determined the cost-effectiveness of AV based on a decision-tree model from a public pntd.0004568 payer perspective. Specific AVs included in the model were Antivipmyn, FAV Afrique, Echi- Editor: H Janaka de Silva, University of Kelaniya, Tab-G and EchiTab-Plus. We derived inputs from the literature which included: type of SRI LANKA snakes causing bites (carpet viper (Echis species)/non-carpet viper), AV effectiveness Received: October 19, 2015 against death, mortality without AV, probability of Early Adverse Reactions (EAR), likelihood Accepted: March 2, 2016 of death from EAR, average age at envenomation in years, anticipated remaining life span Published: March 30, 2016 and likelihood of amputation.