Howell Final Pages.Indb 1 9/22/11 4:58 PM Copyrighted Material Introduction
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Galápagos Natural History Extravaganza
GALÁPAGOS NATURAL HISTORY EXTRAVAGANZA 21 NOVEMBER – 02 DECEMBER 2021 20 NOVEMBER – 01 DECEMBER 2022 19 – 30 NOVEMBER 2023 Red-footed Booby is one of three booby species likely to be found on this trip. www.birdingecotours.com [email protected] 2 | ITINERARY Galapagos: Natural History Extravaganza Just 483 years ago the first man stepped onto the Galápagos Islands and marveled at this living laboratory. Today we continue to be awestruck by this constantly changing archipelago. As the Nazca Plate moves and new islands are formed, evolution is illustrated up close and personal. Visiting the Galápagos archipelago is a dream for all naturalists. From Quito we will fly to the island of Baltra. We then will make our way to our home for the next eight days, the Samba, a spacious and luxuriously designed motor sailing yacht with wide open and shaded sun decks, a fully stocked bar, and a library. The abundant marine life that visits these waters year-round, the Marine Iguanas that rule the rocky coastlines, and of course a unique group of birds make it easy to understand why this trip is a must for birders and natural history buffs. Some of the Galápagos specials that we hope to find on this trip include Galapagos Penguin, Waved Albatross, Galapagos Shearwater, Wedge-rumped and Elliot’s Storm Petrels, Magnificent Frigatebird, Nazca, Red-footed, and Blue-footed Boobies, Lava Heron, Galapagos Hawk, Lava Gull, Galapagos Martin, Galapagos Flycatcher, Vermilion Flycatcher, Galapagos, Floreana, San Cristobal, and Espanola Mockingbirds, Vegetarian Finch, Woodpecker Finch, Common Cactus Finch, Green Warbler-Finch, Large and Small Tree Finches, Small and Medium Ground Finches, and Mangrove Finch. -

Ecuador's Biodiversity Hotspots
Ecuador’s Biodiversity Hotspots Destination: Andes, Amazon & Galapagos Islands, Ecuador Duration: 19 Days Dates: 29th June – 17th July 2018 Exploring various habitats throughout the wonderful & diverse country of Ecuador Spotting a huge male Andean bear & watching as it ripped into & fed on bromeliads Watching a Eastern olingo climbing the cecropia from the decking in Wildsumaco Seeing ~200 species of bird including 33 species of dazzling hummingbirds Watching a Western Galapagos racer hunting, catching & eating a Marine iguana Incredible animals in the Galapagos including nesting flightless cormorants 36 mammal species including Lowland paca, Andean bear & Galapagos fur seals Watching the incredible and tiny Pygmy marmoset in the Amazon near Sacha Lodge Having very close views of 8 different Andean condors including 3 on the ground Having Galapagos sea lions come up & interact with us on the boat and snorkelling Tour Leader / Guides Overview Martin Royle (Royle Safaris Tour Leader) Gustavo (Andean Naturalist Guide) Day 1: Quito / Puembo Francisco (Antisana Reserve Guide) Milton (Cayambe Coca National Park Guide) ‘Campion’ (Wildsumaco Guide) Day 2: Antisana Wilmar (Shanshu), Alex and Erica (Amazonia Guides) Gustavo (Galapagos Islands Guide) Days 3-4: Cayambe Coca Participants Mr. Joe Boyer Days 5-6: Wildsumaco Mrs. Rhoda Boyer-Perkins Day 7: Quito / Puembo Days 8-10: Amazon Day 11: Quito / Puembo Days 12-18: Galapagos Day 19: Quito / Puembo Royle Safaris – 6 Greenhythe Rd, Heald Green, Cheshire, SK8 3NS – 0845 226 8259 – [email protected] Day by Day Breakdown Overview Ecuador may be a small country on a map, but it is one of the richest countries in the world in terms of life and biodiversity. -

Lista Oficial PN
Nombre científico Inglés Español Crypturellus souiC Little Tinamou Tinamú Chico Crypturellus cinnamomeusR Thicket Tinamou Tinamú Canelo Dendrocygna autumnalis Black-bellied Whistling-Duck Pijije Común Cairina moschata Muscovy Duck Pato Real Ortalis vetula Plain Chachalaca Chachalaca Olivácea Penelope purpurascens Crested Guan Pava Crestada Crax rubra Great Curassow Pavón Grande Colinus cristatus Crested Bobwhite Codorniz Crestada Tachybaptus dominicus Least Grebe Zambullidor Enano Ardenna creatopus Pink-footed Shearwater Pardela Blanca Común Ardenna pacificus Wedge-tailed Shearwater Pardela Colicuña Ardenna grisea Sooty Shearwater Pardela Sombría Puffinus nativitatis Christmas Shearwater Pardela de Navidad, Pardela de Christmas Puffinus subalaris Galapagos Shearwater Pardela de las Galápagos Puffinus opisthomelas Black-vented Shearwater Pardela Culinegra Oceanodroma leucorhoa Leach's Storm-Petrel Paiño de Leach Oceanodroma tethys Wedge-rumped Storm-Petrel Paiño Danzarin Oceanodroma melania Black Storm-Petrel Paiño Negro Oceanodroma microsoma Least Storm-Petrel Paiño Menudo Phaethon aethereus Red-billed Tropicbird Rabijunco Piquirrojo Mycteria americana Wood Stork Cigüeñón Fregata magnificens Magnificent Frigatebird Rabihorcado Magno Sula dactylatra Masked Booby Piquero Blanco Sula granti Nazca Booby Piquero de Nazca AOCR Sula nebouxiiC Blue-footed Booby Piquero Patiazul Sula variegta Peruvian Booby Piquero Peruano Sula leucogasterR Brown Booby Piquero Moreno Sula sula Red-footed Booby Piquero Patirrojo Phalacrocorax brasilianus Neotropic -

Observations on the Breeding and Distribution of Lava Gull Leucophaeus Fuliginosus K
Cotinga 37 Observations on the breeding and distribution of Lava Gull Leucophaeus fuliginosus K. Thalia Grant, Olivia H. Estes and Gregory B. Estes Received 7 January 2014; final revision accepted 3 December 2014 Cotinga 37 (2015): OL 22–37 published online 10 March 2015 La Gaviota de Lava Leucophaeus fuliginosus, endémica del archipiélago de Galápagos, es la gaviota más rara del mundo, cuyos hábitos de reproducción son poco conocidos. En los años 2011 y 2012 se observó anidación en la isla Genovesa en densidades mayores a las reportadas previamente en Galápagos. Las parejas reproductoras fueron muy territoriales, defendiendo áreas de hasta 70 m de diámetro contra coespecíficos. Las hembras fueron más agresivas que los machos frente a los intrusos de otras especias percibidos como una amenaza. La nidada de 1–2 huevos fue incubada por ambos miembros de la pareja en turnos de dos horas. Los polluelos salieron del nido 4–5 días después de la eclosión, seleccionando lugares en el territorio más protegidos a los cuales retornaron regularmente para descansar. Los adultos reproductores fueron depredadores oportunistas, alimentando a sus crías principalmente con huevos y polluelos de aves marinas y peces robados de las mismas aves. Reconocemos una relación parasítica entre la cleptoparásita Fragata Real Fregata magnificens y la Gaviota de Lava, y sospechamos que este es el medio principal por el cual en esta isla las gaviotas adquieren los peces que comen. Presentamos datos sobre las comunicaciones entre los padres y la cría, proporcionamos la primera serie de fotografías del desarrollo del polluelo de Gaviota de Lava y describimos una característica en el plumaje de los adultos que no ha sido descrita previamente. -

Alpha Codes for 2168 Bird Species (And 113 Non-Species Taxa) in Accordance with the 62Nd AOU Supplement (2021), Sorted Taxonomically
Four-letter (English Name) and Six-letter (Scientific Name) Alpha Codes for 2168 Bird Species (and 113 Non-Species Taxa) in accordance with the 62nd AOU Supplement (2021), sorted taxonomically Prepared by Peter Pyle and David F. DeSante The Institute for Bird Populations www.birdpop.org ENGLISH NAME 4-LETTER CODE SCIENTIFIC NAME 6-LETTER CODE Highland Tinamou HITI Nothocercus bonapartei NOTBON Great Tinamou GRTI Tinamus major TINMAJ Little Tinamou LITI Crypturellus soui CRYSOU Thicket Tinamou THTI Crypturellus cinnamomeus CRYCIN Slaty-breasted Tinamou SBTI Crypturellus boucardi CRYBOU Choco Tinamou CHTI Crypturellus kerriae CRYKER White-faced Whistling-Duck WFWD Dendrocygna viduata DENVID Black-bellied Whistling-Duck BBWD Dendrocygna autumnalis DENAUT West Indian Whistling-Duck WIWD Dendrocygna arborea DENARB Fulvous Whistling-Duck FUWD Dendrocygna bicolor DENBIC Emperor Goose EMGO Anser canagicus ANSCAN Snow Goose SNGO Anser caerulescens ANSCAE + Lesser Snow Goose White-morph LSGW Anser caerulescens caerulescens ANSCCA + Lesser Snow Goose Intermediate-morph LSGI Anser caerulescens caerulescens ANSCCA + Lesser Snow Goose Blue-morph LSGB Anser caerulescens caerulescens ANSCCA + Greater Snow Goose White-morph GSGW Anser caerulescens atlantica ANSCAT + Greater Snow Goose Intermediate-morph GSGI Anser caerulescens atlantica ANSCAT + Greater Snow Goose Blue-morph GSGB Anser caerulescens atlantica ANSCAT + Snow X Ross's Goose Hybrid SRGH Anser caerulescens x rossii ANSCAR + Snow/Ross's Goose SRGO Anser caerulescens/rossii ANSCRO Ross's Goose -

Trip Report February 24 – March 3, 2019 |Written by Susan and Burt Mittelstadt
Journey to the Galápagos | Trip Report February 24 – March 3, 2019 |Written by Susan and Burt Mittelstadt With Hosts: Susan and Burt Mittelstadt and 12 participants: Mariel, Roy, Jill, Maggie, Baird, Janis, Chuck, Mary, Peggy, Bobbie, Julia and Janet. Saturday, Nov. 23 | Antisana National Park Before flying to the Galápagos, our group of fourteen travelers took an excursion to Antisana National Park to see creatures of the high Andes. Rain was predicted for the afternoon, so we got an early start despite some late-night arrivals. Despite the predictions, it was as clear a day as one could hope in the mountains. Manuel was our knowledgeable guide and Jonathon our expert driver: our goal was the Andean Condor. At the first pullout it was clear we weren’t at sea level. Despite pounding hearts and fuzzy heads, (the elevation all morning was near or over 13,000’) we were all thrilled with instant success. Andean Condors were spotted by Maggie, at rest on cliffs across the deep canyon. Manuel set up his spotting scope and showed us how to take great photos with our phones and the scope. The day continued to amaze. Condors were spotted at a total of three locations with one very close flyby. Lunch Naturalist Journeys, LLC | Caligo Ventures PO Box 16545 Portal, AZ 85632 PH: 520.558.1146 | 866.900.1146 Fax 650.471.7667 naturalistjourneys.com | caligo.com [email protected] | [email protected] was a delicious Andean feast. After lunch, the rain started to come down just as we pulled away to head for our hotel. -

Species List
Naturalist Journeys and Caligo Ventures Galapagos Birding and Wildlife Tour Species List – May 29 – June 5, 2017 With note from Ecuador (Puembo/Antisana National Park 1 day-pre-trip) Guides Carol Simon & Howard Topoff, with local experts and 18 participants Galapagos Islands Birds (40 species): Anatidae: Ducks, Geese, and Swans (1) White-cheeked Pintail Anas bahamensis— seen Tuesday on Floreana Phoenicopteridae: Flamingos (1) American Flamingo Phoenicopterus ruber— Several in brackish lagoon at Punta Cormorant, Floreana Island seen on Wed Spheniscidae: Penguins (1) Galapagos Penguin Spheniscus mendiculus— seen near Punta Cormorant Wed; Bartolome on Fri Procellariidae: Petrels and Shearwaters (1) Galapagos Shearwater Puffinus subalaris— Open water bird seen near San Cristobal Mon/ Espanola Tues/ Floreana Wed / South Plazas Sat Hydrobatidae: Storm-Petrels (1) Elliot’s Storm Petrel Oceanites gracilis— Common throughout Galapagos, skittering across the water Phaethontidae: Tropicbirds (1) Red-billed Tropicbird Phaethon aethereus— Tues flying near Espanola/on nest off Champion Island near Florana Wed/ South Plazas Sat Fregatidae: Frigatebirds (2) Magnificent Frigatebird Fregata magnificens— Seen toward beginning, San Cristobal, Sun/ following our boat on Mon / Off Espanola Tues / Floreana on Wednesday . Santa Cruz Thursday /off Bartolome Fri / North Seymour Sat / on the boat Sun Great Frigatebird Fregata minor— on Punta Pitt off San Cristobal Monday / on North Seymour Sat Sulidae: Gannets and Boobies (3) Nazca Booby Sula granti— Seen on bridge -

Galapagos Endemics Cruise: October-November 2013
Tropical Birding Trip Report Galapagos Endemics Cruise: October-November 2013 A Tropical Birding SET DEPARTURE tour Galapagos Endemics Cruise 24 October – 2 November 2013 A visit to the largest Waved Albatross colony in the world was an undoubted highlight Tropical Birding Tour leader: Jose Illanes (With Galapagos Naturalist Guide Jairo) 1 www.tropicalbirding.com +1-409-515-0514 [email protected] Page Tropical Birding Trip Report Galapagos Endemics Cruise: October-November 2013 Itinerary: Day 1 Arrival in Quito/night Quito Day 2 Quito to Baltra and Santa Cruz Day 3 Genovesa Day 4 Isabela to Bartolome Day 5 Bartolome to San Cristobal Day 6 San Cristobal to Española Day 7 Española Day 8 Floreana and Santa Cruz Day 9 North Seymour to Quito Day 10 Departure from Quito October 25: Quito to Baltra and Santa Cruz As for anybody going to the Galapagos Islands, it is perceived, correctly, as unique and amazing, and, as we were sitting on the plane, everybody’s minds were focused on getting there, and wishing they were there already! I had not been to the Galapagos for a while, and so I had my own excitement of returning to the islands myself knowing this would be a marked change from my usual tours within the jungles and rainforests of the mainland. This was to be a tour where the views of the birds were my least concern, as everything shows well. As we arrived at the main entry port to the Galapagos, the tiny island of Baltra, 2 www.tropicalbirding.com +1-409-515-0514 [email protected] Page Tropical Birding Trip Report Galapagos Endemics Cruise: October-November 2013 everyone’s eyes were glued to the window, taking in their first sights of the islands, which, for some, included their first Galapagos Doves flying by. -
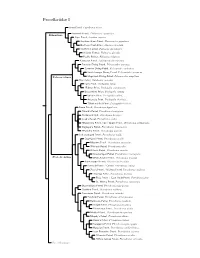
Procellariidae Species Tree
Procellariidae I Snow Petrel, Pagodroma nivea Antarctic Petrel, Thalassoica antarctica Fulmarinae Cape Petrel, Daption capense Southern Giant-Petrel, Macronectes giganteus Northern Giant-Petrel, Macronectes halli Southern Fulmar, Fulmarus glacialoides Atlantic Fulmar, Fulmarus glacialis Pacific Fulmar, Fulmarus rodgersii Kerguelen Petrel, Aphrodroma brevirostris Peruvian Diving-Petrel, Pelecanoides garnotii Common Diving-Petrel, Pelecanoides urinatrix South Georgia Diving-Petrel, Pelecanoides georgicus Pelecanoidinae Magellanic Diving-Petrel, Pelecanoides magellani Blue Petrel, Halobaena caerulea Fairy Prion, Pachyptila turtur ?Fulmar Prion, Pachyptila crassirostris Broad-billed Prion, Pachyptila vittata Salvin’s Prion, Pachyptila salvini Antarctic Prion, Pachyptila desolata ?Slender-billed Prion, Pachyptila belcheri Bonin Petrel, Pterodroma hypoleuca ?Gould’s Petrel, Pterodroma leucoptera ?Collared Petrel, Pterodroma brevipes Cook’s Petrel, Pterodroma cookii ?Masatierra Petrel / De Filippi’s Petrel, Pterodroma defilippiana Stejneger’s Petrel, Pterodroma longirostris ?Pycroft’s Petrel, Pterodroma pycrofti Soft-plumaged Petrel, Pterodroma mollis Gray-faced Petrel, Pterodroma gouldi Magenta Petrel, Pterodroma magentae ?Phoenix Petrel, Pterodroma alba Atlantic Petrel, Pterodroma incerta Great-winged Petrel, Pterodroma macroptera Pterodrominae White-headed Petrel, Pterodroma lessonii Black-capped Petrel, Pterodroma hasitata Bermuda Petrel / Cahow, Pterodroma cahow Zino’s Petrel / Madeira Petrel, Pterodroma madeira Desertas Petrel, Pterodroma -

Shearwatersrefs V1.10.Pdf
Introduction I have endeavoured to keep typos, errors, omissions etc in this list to a minimum, however when you find more I would be grateful if you could mail the details during 2018 & 2019 to: [email protected]. Please note that this and other Reference Lists I have compiled are not exhaustive and are best employed in conjunction with other sources. Grateful thanks to Ashley Fisher (www.scillypelagics.com) for the cover images. All images © the photographer. Joe Hobbs Index The general order of species follows the International Ornithologists' Union World Bird List (Gill, F. & Donsker, D. (eds.) 2017. IOC World Bird List. Available from: http://www.worldbirdnames.org/ [version 7.3 accessed August 2017]). Version Version 1.10 (January 2018). Cover Main image: Great Shearwater. At sea 3’ SW of the Bishop Rock, Isles of Scilly. 8th August 2009. Picture by Ashley Fisher. Vignette: Sooty Shearwater. At sea off the Isles of Scilly. 14th August 2009. Picture by Ashley Fisher. Species Page No. Audubon's Shearwater [Puffinus lherminieri] 34 Balearic Shearwater [Puffinus mauretanicus] 28 Bannerman's Shearwater [Puffinus bannermani] 37 Barolo Shearwater [Puffinus baroli] 38 Black-vented Shearwater [Puffinus opisthomelas] 30 Boyd's Shearwater [Puffinus boydi] 38 Bryan's Shearwater [Puffinus bryani] 29 Buller's Shearwater [Ardenna bulleri] 15 Cape Verde Shearwater [Calonectris edwardsii] 12 Christmas Island Shearwater [Puffinus nativitatis] 23 Cory's Shearwater [Calonectris borealis] 9 Flesh-footed Shearwater [Ardenna carneipes] 21 Fluttering -

Threats to Seabirds: a Global Assessment 2 3 4 Authors: Maria P
1 Threats to seabirds: a global assessment 2 3 4 Authors: Maria P. Dias1*, Rob Martin1, Elizabeth J. Pearmain1, Ian J. Burfield1, Cleo Small2, Richard A. 5 Phillips3, Oliver Yates4, Ben Lascelles1, Pablo Garcia Borboroglu5, John P. Croxall1 6 7 8 Affiliations: 9 1 - BirdLife International. The David Attenborough Building, Pembroke Street Cambridge CB2 3QZ UK 10 2 - BirdLife International Marine Programme, RSPB, The Lodge, Sandy, SG19 2DL 11 3 – British Antarctic Survey. Natural Environment Research Council, High Cross, Madingley Road, 12 Cambridge CB3 0ET, UK 13 4 – Centre for the Environment, Fishery and Aquaculture Science, Pakefield Road, Lowestoft, NR33, UK 14 5 - Global Penguin Society, University of Washington and CONICET Argentina. Puerto Madryn U9120, 15 Chubut, Argentina 16 * Corresponding author: Maria Dias, [email protected]. BirdLife International. The David 17 Attenborough Building, Pembroke Street Cambridge CB2 3QZ UK. Phone: +44 (0)1223 747540 18 19 20 Acknowledgements 21 We are very grateful to Bartek Arendarczyk, Sophie Bennett, Ricky Hibble, Eleanor Miller and Amy 22 Palmer-Newton for assisting with the bibliographic review. We thank Rachael Alderman, Pep Arcos, 23 Jonathon Barrington, Igor Debski, Peter Hodum, Gustavo Jimenez, Jeff Mangel, Ken Morgan, Paul Sagar, 24 Peter Ryan, and other members of the ACAP PaCSWG, and the members of IUCN SSC Penguin Specialist 25 Group (Alejandro Simeone, Andre Chiaradia, Barbara Wienecke, Charles-André Bost, Lauren Waller, Phil 26 Trathan, Philip Seddon, Susie Ellis, Tom Schneider and Dee Boersma) for reviewing threats to selected 27 species. We thank also Andy Symes, Rocio Moreno, Stuart Butchart, Paul Donald, Rory Crawford, 28 Tammy Davies, Ana Carneiro and Tris Allinson for fruitful discussions and helpful comments on earlier 29 versions of the manuscript. -

Pacific Seabird Group Committee Coordinators
PACIFIC SEABIRDS A Publication of the Pacific Seabird Group Volume 39, Number 1 Spring 2012 PACIFIC SEABIRD GROUP Dedicated to the Study and Conservation of Pacific Seabirds and Their Environment The Pacific Seabird Group (PSG) was formed in 1972 due to the need for better communication among Pacific seabird researchers. PSG provides a forum for the research activities of its members, promotes the conservation of seabirds, and informs members and the public of issues relating to Pacific Ocean seabirds and their environment. PSG members include research scientists, conservation professionals, and members of the public from all parts of the Pacific Ocean. The group also welcomes seabird professionals and enthusiasts in other parts of the world. PSG holds annual meetings at which scientific papers and symposia are presented; abstracts for meetings are published on our web site. The group is active in promoting conservation of seabirds, include seabird/fisheries interactions, monitoring of seabird populations, seabird restoration following oil spills, establishment of seabird sanctuaries, and endangered species. Policy statements are issued on conservation issues of critical importance. PSG’s journals are Pacific Seabirds (formerly the PSG Bulletin) and Marine Ornithology. Other publications include symposium volumes and technical reports; these are listed near the back of this issue. PSG is a member of the International Union for Conservation of Nature (IUCN), the Ornithological Council, and the American Bird Conservancy. Annual dues for membership are $30 (individual and family); $24 (student, undergraduate and graduate); and $900 (Life Membership, payable in five $180 installments). Dues are payable to the Treasurer; see the PSG web site, or the Membership Order Form next to inside back cover.