The Genus Fulgoraria (Gastropoda: Volutidae)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

(Approx) Mixed Micro Shells (22G Bags) Philippines € 10,00 £8,64 $11,69 Each 22G Bag Provides Hours of Fun; Some Interesting Foraminifera Also Included
Special Price £ US$ Family Genus, species Country Quality Size Remarks w/o Photo Date added Category characteristic (€) (approx) (approx) Mixed micro shells (22g bags) Philippines € 10,00 £8,64 $11,69 Each 22g bag provides hours of fun; some interesting Foraminifera also included. 17/06/21 Mixed micro shells Ischnochitonidae Callistochiton pulchrior Panama F+++ 89mm € 1,80 £1,55 $2,10 21/12/16 Polyplacophora Ischnochitonidae Chaetopleura lurida Panama F+++ 2022mm € 3,00 £2,59 $3,51 Hairy girdles, beautifully preserved. Web 24/12/16 Polyplacophora Ischnochitonidae Ischnochiton textilis South Africa F+++ 30mm+ € 4,00 £3,45 $4,68 30/04/21 Polyplacophora Ischnochitonidae Ischnochiton textilis South Africa F+++ 27.9mm € 2,80 £2,42 $3,27 30/04/21 Polyplacophora Ischnochitonidae Stenoplax limaciformis Panama F+++ 16mm+ € 6,50 £5,61 $7,60 Uncommon. 24/12/16 Polyplacophora Chitonidae Acanthopleura gemmata Philippines F+++ 25mm+ € 2,50 £2,16 $2,92 Hairy margins, beautifully preserved. 04/08/17 Polyplacophora Chitonidae Acanthopleura gemmata Australia F+++ 25mm+ € 2,60 £2,25 $3,04 02/06/18 Polyplacophora Chitonidae Acanthopleura granulata Panama F+++ 41mm+ € 4,00 £3,45 $4,68 West Indian 'fuzzy' chiton. Web 24/12/16 Polyplacophora Chitonidae Acanthopleura granulata Panama F+++ 32mm+ € 3,00 £2,59 $3,51 West Indian 'fuzzy' chiton. 24/12/16 Polyplacophora Chitonidae Chiton tuberculatus Panama F+++ 44mm+ € 5,00 £4,32 $5,85 Caribbean. 24/12/16 Polyplacophora Chitonidae Chiton tuberculatus Panama F++ 35mm € 2,50 £2,16 $2,92 Caribbean. 24/12/16 Polyplacophora Chitonidae Chiton tuberculatus Panama F+++ 29mm+ € 3,00 £2,59 $3,51 Caribbean. -

Miocene Vetigastropoda and Neritimorpha (Mollusca, Gastropoda) of Central Chile
Journal of South American Earth Sciences 17 (2004) 73–88 www.elsevier.com/locate/jsames Miocene Vetigastropoda and Neritimorpha (Mollusca, Gastropoda) of central Chile Sven N. Nielsena,*, Daniel Frassinettib, Klaus Bandela aGeologisch-Pala¨ontologisches Institut und Museum, Universita¨t Hamburg, Bundesstrasse 55, 20146 Hamburg, Germany bMuseo Nacional de Historia Natural, Casilla 787, Santiago, Chile Abstract Species of Vetigastropoda (Fissurellidae, Turbinidae, Trochidae) and one species of Neritimorpha (Neritidae) from the Navidad area, south of Valparaı´so, and the Arauco Peninsula, south of Concepcio´n, are described. Among these, the Fissurellidae comprise Diodora fragilis n. sp., Diodora pupuyana n. sp., two additional unnamed species of Diodora, and a species resembling Fissurellidea. Turbinidae are represented by Cantrainea sp., and Trochidae include Tegula (Chlorostoma) austropacifica n. sp., Tegula (Chlorostoma) chilena n. sp., Tegula (Chlorostoma) matanzensis n. sp., Tegula (Agathistoma) antiqua n. sp., Bathybembix mcleani n. sp., Gibbula poeppigii [Philippi, 1887] n. comb., Diloma miocenica n. sp., Fagnastesia venefica [Philippi, 1887] n. gen. n. comb., Fagnastesia matanzana n. gen. n. sp., Calliostoma mapucherum n. sp., Calliostoma kleppi n. sp., Calliostoma covacevichi n. sp., Astele laevis [Sowerby, 1846] n. comb., and Monilea riorapelensis n. sp. The Neritidae are represented by Nerita (Heminerita) chilensis [Philippi, 1887]. The new genus Fagnastesia is introduced to represent low-spired trochoideans with a sculpture of nodes below the suture, angulated whorls, and a wide umbilicus. This Miocene Chilean fauna includes genera that have lived at the coast and in shallow, relatively warm water or deeper, much cooler water. This composition therefore suggests that many of the Miocene formations along the central Chilean coast consist of displaced sediments. -

Anatomy of Zetela Alphonsi Vilvens, 2002 Casts Doubt on Its Original Placement Based on Conchological Characters
SPIXIANA 40 2 161-170 München, Dezember 2017 ISSN 0341-8391 Anatomy of Zetela alphonsi Vilvens, 2002 casts doubt on its original placement based on conchological characters (Mollusca, Solariellidae) Enrico Schwabe, Martin Heß, Lauren Sumner-Rooney & Javier Sellanes Schwabe, E., Heß, M., Sumner-Rooney, L. H. & Sellanes, J. 2017. Anatomy of Zetela alphonsi Vilvens, 2002 casts doubt on its original placement based on con- chological characters (Mollusca, Solariellidae). Spixiana 40 (2): 161-170. Zetela is a genus of marine gastropods belonging to the family Solariellidae (Vetigastropoda, Trochoidea). The species Zetela alphonsi Vilvens, 2002 was origi- nally described using classical conchological characters from samples obtained from deep water off Chiloé, Chile, but no other features were recorded in the original description. In the present paper, taxonomically relevant characters such as the operculum, radula, and soft part anatomy are illustrated and described for the first time from a specimen of Zetela alphonsi from a new locality, a methane seep area off the coast of central Chile. We find that many features differ substantially from other members of the genus, including several which deviate from characters that are diagnostic for Zetela. Zetela alphonsi also lacks retinal screening pigment, giving the appearance of eyelessness from gross examination. Although pigmenta- tion loss has been reported in other deep sea genera, this feature has not been re- ported in any other members of Zetela. We reconstructed a tomographic model of the eye and eyestalk, and demonstrate that a vestigial eye is still present but exter- nally invisible due to the loss of pigmentation. Based on these new morphological characters, particularly observations of the radula, we suggest that the current placement of this species in the genus Zetela should be reconsidered. -

An Annotated Checklist of the Marine Macroinvertebrates of Alaska David T
NOAA Professional Paper NMFS 19 An annotated checklist of the marine macroinvertebrates of Alaska David T. Drumm • Katherine P. Maslenikov Robert Van Syoc • James W. Orr • Robert R. Lauth Duane E. Stevenson • Theodore W. Pietsch November 2016 U.S. Department of Commerce NOAA Professional Penny Pritzker Secretary of Commerce National Oceanic Papers NMFS and Atmospheric Administration Kathryn D. Sullivan Scientific Editor* Administrator Richard Langton National Marine National Marine Fisheries Service Fisheries Service Northeast Fisheries Science Center Maine Field Station Eileen Sobeck 17 Godfrey Drive, Suite 1 Assistant Administrator Orono, Maine 04473 for Fisheries Associate Editor Kathryn Dennis National Marine Fisheries Service Office of Science and Technology Economics and Social Analysis Division 1845 Wasp Blvd., Bldg. 178 Honolulu, Hawaii 96818 Managing Editor Shelley Arenas National Marine Fisheries Service Scientific Publications Office 7600 Sand Point Way NE Seattle, Washington 98115 Editorial Committee Ann C. Matarese National Marine Fisheries Service James W. Orr National Marine Fisheries Service The NOAA Professional Paper NMFS (ISSN 1931-4590) series is pub- lished by the Scientific Publications Of- *Bruce Mundy (PIFSC) was Scientific Editor during the fice, National Marine Fisheries Service, scientific editing and preparation of this report. NOAA, 7600 Sand Point Way NE, Seattle, WA 98115. The Secretary of Commerce has The NOAA Professional Paper NMFS series carries peer-reviewed, lengthy original determined that the publication of research reports, taxonomic keys, species synopses, flora and fauna studies, and data- this series is necessary in the transac- intensive reports on investigations in fishery science, engineering, and economics. tion of the public business required by law of this Department. -
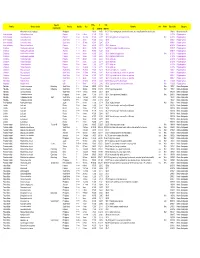
Approx) (Approx) Mixed Micro Shells (22G Bags) Philippines € 10,00 £8,56 $11,71 Each 22G Bag Provides Hours of Fun; Some Interesting Foraminifera Also Included
Special Price £ US$ Family Genus, species Country Quality Size Remarks w/o Photo Date added Category characteristic (€) (approx) (approx) Mixed micro shells (22g bags) Philippines € 10,00 £8,56 $11,71 Each 22g bag provides hours of fun; some interesting Foraminifera also included. 17/06/21 Mixed micro shells Ischnochitonidae Callistochiton pulchrior Panama F+++ 89mm € 1,80 £1,54 $2,11 21/12/16 Polyplacophora Ischnochitonidae Chaetopleura lurida Panama F+++ 2022mm € 3,00 £2,57 $3,51 Hairy girdles, beautifully preserved. Web 24/12/16 Polyplacophora Ischnochitonidae Ischnochiton textilis South Africa F+++ 30mm+ € 4,00 £3,42 $4,68 30/04/21 Polyplacophora Ischnochitonidae Ischnochiton textilis South Africa F+++ 27.9mm € 2,80 £2,40 $3,28 30/04/21 Polyplacophora Ischnochitonidae Stenoplax limaciformis Panama F+++ 16mm+ € 6,50 £5,57 $7,61 Uncommon. 24/12/16 Polyplacophora Chitonidae Acanthopleura gemmata Philippines F+++ 25mm+ € 2,50 £2,14 $2,93 Hairy margins, beautifully preserved. 04/08/17 Polyplacophora Chitonidae Acanthopleura gemmata Australia F+++ 25mm+ € 2,60 £2,23 $3,05 02/06/18 Polyplacophora Chitonidae Acanthopleura granulata Panama F+++ 41mm+ € 4,00 £3,42 $4,68 West Indian 'fuzzy' chiton. Web 24/12/16 Polyplacophora Chitonidae Acanthopleura granulata Panama F+++ 32mm+ € 3,00 £2,57 $3,51 West Indian 'fuzzy' chiton. 24/12/16 Polyplacophora Chitonidae Chiton tuberculatus Panama F+++ 44mm+ € 5,00 £4,28 $5,86 Caribbean. 24/12/16 Polyplacophora Chitonidae Chiton tuberculatus Panama F++ 35mm € 2,50 £2,14 $2,93 Caribbean. 24/12/16 Polyplacophora Chitonidae Chiton tuberculatus Panama F+++ 29mm+ € 3,00 £2,57 $3,51 Caribbean. -

Evolutionary Patterns and Consequences of Developmental Mode in Cenozoic Gastropods from Southeastern Australia
Evolutionary patterns and consequences of developmental mode in Cenozoic gastropods from southeastern Australia Thesis submitted in accordance with the requirements of the University of Liverpool for the degree of Doctor in Philosophy by Kirstie Rae Thomson September 2013 ABSTRACT Gastropods, like many other marine invertebrates undergo a two-stage life cycle. As the adult body plan results in narrow environmental tolerances and restricted mobility, the optimum opportunity for dispersal occurs during the initial larval phase. Dispersal is considered to be a major influence on the evolutionary trends of different larval strategies. Three larval strategies are recognised in this research: planktotrophy, lecithotrophy and direct development. Planktotrophic larvae are able to feed and swim in the plankton resulting in the greatest dispersal potential. Lecithotrophic larvae have a reduced planktic period and are considered to have more restricted dispersal. The planktic period is absent in direct developing larvae and therefore dispersal potential in these taxa is extremely limited. Each of these larval strategies can be confidently inferred from the shells of fossil gastropods and the evolutionary trends associated with modes of development can be examined using both phylogenetic and non-phylogenetic techniques. This research uses Cenozoic gastropods from southeastern Australia to examine evolutionary trends associated with larval mode. To ensure the species used in analyses are distinct and correctly assigned, a taxonomic review of the six families included in this study was undertaken. The families included in this study were the Volutidae, Nassariidae, Raphitomidae, Borsoniidae, Mangeliidae and Turridae. Phylogenetic analyses were used to examine the relationships between taxa and to determine the order and timing of changes in larval mode throughout the Cenozoic. -

James Hamilton Mclean: the Master of the Gastropoda
Zoosymposia 13: 014–043 (2019) ISSN 1178-9905 (print edition) http://www.mapress.com/j/zs/ ZOOSYMPOSIA Copyright © 2019 · Magnolia Press ISSN 1178-9913 (online edition) http://dx.doi.org/10.11646/zoosymposia.13.1.4 http://zoobank.org/urn:lsid:zoobank.org:pub:20E93C08-5C32-42FC-9580-1DED748FCB5F James Hamilton McLean: The master of the Gastropoda LINDSEY T. GROVES1, DANIEL L. GEIGER2, JANN E. VENDETTI1, & EUGENE V. COAN3 1Natural History Museum of Los Angeles County, Malacology Department, 900 Exposition Blvd., Los Angeles, California 90007, U.S.A. E-mail: [email protected]; [email protected] 2Santa Barbara Museum of Natural History, Department of Invertebrate Zoology, 2559 Puesta del Sol, Santa Barbara, California 93105, U.S.A. E-mail: [email protected] 3P.O. Box 420495, Summerland Key, Florida 33042, U.S.A. E-mail: [email protected] Abstract A biography of the late James H. McLean, former Curator of Malacology at the Natural History Museum of Los Angeles County is provided. It is complemented with a full bibliography and list of 344 taxa named by him and co-authors (with type information and current status), as well as 40 patronyms. Biography James Hamilton McLean was born in Detroit, Michigan, on June 17, 1936. The McLean family moved to Dobbs Ferry, New York, on the Hudson River in 1940, a short train ride and subway ride away from the American Museum of Natural History (AMNH). His brother Hugh recalled that, “AMNH became the place of choice to go to whenever we could get someone to take us. Those visits opened our eyes to the variety and possibilities of what was out there, waiting for us to discover and collect.” From an early age James seemed destined to have a career at a museum (Figs 1–2). -

Miocene Shallow Marine Molluscs from the Hokutan Group in the Tajima Area, Hyôgo Prefecture, Southwest Japan
Bulletin of the Mizunami Fossil Museum, no. 37, p. 51–113, 9 pls., 2 fi gs., 1 table. © 2011, Mizunami Fossil Museum Miocene shallow marine molluscs from the Hokutan Group in the Tajima area, Hyôgo Prefecture, southwest Japan Takashi Matsubara Division of Natural History, Museum of Nature and Human Activities, Hyogo, 6 Yayoigaoka, Sanda 669-1546, Japan <[email protected]> Abstract A taxonomical study of molluscs from the Miocene Hokutan Group was carried out and their paleobiogeographic implication was discussed. A total of 100 species, including 45 species of Gastropoda, 54 species of Bivalvia, and one species of Scaphopoda, were discriminated. The fauna is referred to the Kadonosawa Fauna s.s. on the basis of the occurrences of the intertidal “Arcid-Potamid Fauna” and the upper sublittoral Dosinia-Anadara Assemblage. The upper sublittoral assemblages include Turritella (Turritella) kiiensis Yokoyama, Turritella (Hataiella) yoshidai Kotaka, T. (Kurosioia) neiensis Ida, Ostrea sunakozakaensis (Ogasawara), Cucullaea (Cucullaea) toyamaensis Tsuda, Siphonalia osawanoensis Tsuda, and Varicospira toyamaensis (Tsuda). These species are distributed as far north as the southernmost part of northeast Honshû at that time, and their geographic distributions are identical with the Miocene mangrove swamp element. The occurrences of these species strongly support the previous paleoclimatic reconstruction of the Japanese Islands during the latest Early–earliest Middle Miocene age on the basis of the intertidal molluscan assemblages, and show that central and southwest Honshû was under the warmer marine climate than in northeast Honshû and northwards. All the species including a new cardiid, Parvicardium? mikii sp. nov., are described and/or discussed taxonomically. -

T of OCEANOGRAPHY I NENALEM R
/E.ry COL UMB/A HMSC GC 856 .0735 D,cop.2 RTMENT of OCEANOGRAPHY I NENALEM R. TILLAMOOK BAY SCHOOL of SCIENCE W OREGON STATE UNIVERSITY I1I 'Il YAO U/NA ICI ALSEA R. PROGRESS REPORT Ecological Studies of Radioactivity in the Columbia River Estuary and Adjacent Pacific Ocean OREGON STATE UNIVERSITY Wayne V. Burt, Principal Investigator William O. Forster, Research Coordinator Compiled and Edited by James E. McCauley Atomic Energy Commission Contract AT(45-1)1750 MARILYN POTTS GUIN LIBRARY RLO-1750-28 HATFIELD MARINE SCIENCE CENTER QPEGON STATE UNIVERSITY Reference 68-7 It.,, wT, OREGON 97365 1 July 1967 through 30 June 1968 ECOLOGICAL STUDIES OF RADIOACTIVITYIN THE COLUMBIA RIVER ESTUARY AND ADJACENT PACIFICOCEAN Compiled and Edited by James E. McCauley Principal Investigator:Wayne V. Burt Research Coordinator: William O. Forster Co-Investigators: Andrew G. Carey, Jr. James E. McCauley William G. Pearcy William C. Renfro Department of Oceanography Oregon State University Corvallis, Oregon 97331 PROGRESS REPORT 1 July 1967 through 30 June 1968 Submitted to U. S. Atomic Energy Commission Contract AT(45-1)1750 Reference 68-7 RLO 1750-28 July 1968 Iii FE C ,, S,;ENCE CENTER 06LEON STATE 'i!'ERSITY E PQW1, OREGON 97365 STAFF Wayne V. Burt, Ph.D. Principal Investigator Associate Dean of Research Andrew G. Carey, Jr., Ph.D. Co-Investigator Assistant Professor of Oceanography Benthic Ecology William O. Forster, Ph. D. Co-Investigator Assistant Professor of Oceanography Radiochemistry JamesE. McCauley, Ph.D. Co-Investigator Associate Professor of Oceanography Benthic Ecology William G. Pearcy,Ph. D. Co-Investigator Associate Professor of Oceanography Nekton Ecology William C:;Renfro,,Ph.D., Co-Investigator Assistant Professor of Oceanography Radioecology Mary Ann Alspach,, B.A. -

GOA Book.Indb
Elmer E. Rasmuson Library Cataloging in Publication Data The Gulf of Alaska : biology and oceanography / Phillip R. Mundy, ed. – Fairbanks : Alaska Sea Grant College Program, University of Alaska Fairbanks 2005. p. : ill., maps ; cm. – (Alaska Sea Grant College Program ; AK-SG-05-01) Includes bibliographical references and index. ISBN 1-56612-090-x 1. Marine biology—Alaska, Gulf of. 2. Oceanography—Alaska. 3. Ecosystem health—Alaska, Gulf of. I. Title. II. Mundy, Phillip R. (Phillip Roy). Series: Alaska Sea Grant College Program report ; AK-SG-05-01. QH95.35.G845 2005 Citation: Mundy, Phillip R. (ed.). 2005. The Gulf of Alaska: Biology and Oceanography. Alaska Sea Grant College Program, University of Alaska Fairbanks. CREDITS Work for this book was supported by a grant from the Exxon Valdez Oil Spill Trustee Council, Anchorage, Alaska. The Exxon Valdez Oil Spill Trustee Council oversees restoration of the injured ecosystem through a civil settlement that includes the State of Alaska, the U.S. Federal Government, and the Exxon Company. Please see http://www.evostc.state.ak.us. ATMOSP ND HE Publisher of the book is the Alaska Sea Grant College Program, supported by R A IC IC A N D A M E I C N O I S the U.S. Department of Commerce, NOAA National Sea Grant Office, grant L T A R N A T O I I O T N A NA16RG2321, project A/161-01; and by the University of Alaska Fairbanks N U E S C with state funds. The University of Alaska is an affirmative action/equal D R E E P M A M RT O MENT OF C opportunity employer and educational institution. -

Provisional Agenda
CBD Distr. GENERAL UNEP/CBD/SBSTTA/20/INF/24* 11 April 2016 ENGLISH ONLY SUBSIDIARY BODY ON SCIENTIFIC, TECHNICAL AND TECHNOLOGICAL ADVICE Twentieth meeting Montreal, Canada, 25-30 April 2016 Item 4.1 of the provisional agenda** REPORT OF THE REGIONAL WORKSHOP TO FACILITATE THE DESCRIPTION OF ECOLOGICALLY OR BIOLOGICALLY SIGNIFICANT MARINE AREAS IN THE SEAS OF EAST ASIA1 INTRODUCTION 1. At its tenth meeting, the Conference of the Parties to the Convention on Biological Diversity requested the Executive Secretary to work with Parties and other Governments as well as competent organizations and regional initiatives, such as the Food and Agriculture Organization of the United Nations (FAO), regional seas conventions and action plans, and, where appropriate, regional fisheries management organizations (RFMOs) to organize, including the setting of terms of reference, a series of regional workshops, with a primary objective to facilitate the description of ecologically or biologically significant marine areas through the application of scientific criteria in annex I of decision IX/20 as well as other relevant compatible and complementary nationally and intergovernmentally agreed scientific criteria, as well as the scientific guidance on the identification of marine areas beyond national jurisdiction, which meet the scientific criteria in annex I to decision IX/20 (paragraph 36 of decision X/29). 2. In the same decision, the Conference of the Parties requested that the Executive Secretary make available the scientific and technical data, and information and results collated through the workshops referred to above to participating Parties, other Governments, intergovernmental agencies and the Subsidiary Body on Scientific, Technical and Technological Advice (SBSTTA) for their use according to their competencies. -

WCM 2001 Abstract Volume
Abstracts Council of UNITAS MALACOLOGICA 1998-2001 World Congress of President: Luitfried SALVINI-PLAWEN (Wien/Vienna, Austria) Malacology Secretary: Peter B. MORDAN (London, England, UK) Treasurer: Jackie VAN GOETHEM (Bruxelles/Brussels, Belgium) 2001 Members of Council: Takahiro ASAMI (Matsumoto, Japan) Klaus BANDEL (Hamburg, Germany) Yuri KANTOR (Moskwa/Moscow, Russia) Pablo Enrique PENCHASZADEH (Buenos Aires, Argentinia) John D. TAYLOR (London, England, UK) Vienna, Austria Retired President: Rüdiger BIELER (Chicago, USA) 19. – 25. August Edited by Luitfried Salvini-Plawen, Janice Voltzow, Helmut Sattmann and Gerhard Steiner Published by UNITAS MALACOLOGICA, Vienna 2001 I II Organisation of Congress Symposia held at the WCM 2001 Organisers-in-chief: Gerhard STEINER (Universität Wien) Ancient Lakes: Laboratories and Archives of Molluscan Evolution Luitfried SALVINI-PLAWEN (Universität Wien) Organised by Frank WESSELINGH (Leiden, The Netherlands) and Christiane TODT (Universität Wien) Ellinor MICHEL (Amsterdam, The Netherlands) (sponsored by UM). Helmut SATTMANN (Naturhistorisches Museum Wien) Molluscan Chemosymbiosis Organised by Penelope BARNES (Balboa, Panama), Carole HICKMAN Organising Committee (Berkeley, USA) and Martin ZUSCHIN (Wien/Vienna, Austria) Lisa ANGER Anita MORTH (sponsored by UM). Claudia BAUER Rainer MÜLLAN Mathias BRUCKNER Alice OTT Thomas BÜCHINGER Andreas PILAT Hermann DREYER Barbara PIRINGER Evo-Devo in Mollusca Karl EDLINGER (NHM Wien) Heidemarie POLLAK Organised by Gerhard HASZPRUNAR (München/Munich, Germany) Pia Andrea EGGER Eva-Maria PRIBIL-HAMBERGER and Wim J.A.G. DICTUS (Utrecht, The Netherlands) (sponsored by Roman EISENHUT (NHM Wien) AMS). Christine EXNER Emanuel REDL Angelika GRÜNDLER Alexander REISCHÜTZ AMMER CHAEFER Mag. Sabine H Kurt S Claudia HANDL Denise SCHNEIDER Matthias HARZHAUSER (NHM Wien) Elisabeth SINGER Molluscan Conservation & Biodiversity Franz HOCHSTÖGER Mariti STEINER Organised by Ian KILLEEN (Felixtowe, UK) and Mary SEDDON Christoph HÖRWEG Michael URBANEK (Cardiff, UK) (sponsored by UM).