Effect of Chronic Administration of Antidepressants on (12
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
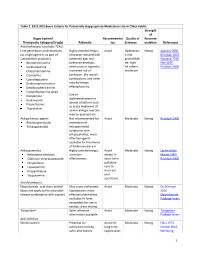
Table 2. 2012 AGS Beers Criteria for Potentially
Table 2. 2012 AGS Beers Criteria for Potentially Inappropriate Medication Use in Older Adults Strength of Organ System/ Recommendat Quality of Recomm Therapeutic Category/Drug(s) Rationale ion Evidence endation References Anticholinergics (excludes TCAs) First-generation antihistamines Highly anticholinergic; Avoid Hydroxyzin Strong Agostini 2001 (as single agent or as part of clearance reduced with e and Boustani 2007 combination products) advanced age, and promethazi Guaiana 2010 Brompheniramine tolerance develops ne: high; Han 2001 Carbinoxamine when used as hypnotic; All others: Rudolph 2008 Chlorpheniramine increased risk of moderate Clemastine confusion, dry mouth, Cyproheptadine constipation, and other Dexbrompheniramine anticholinergic Dexchlorpheniramine effects/toxicity. Diphenhydramine (oral) Doxylamine Use of diphenhydramine in Hydroxyzine special situations such Promethazine as acute treatment of Triprolidine severe allergic reaction may be appropriate. Antiparkinson agents Not recommended for Avoid Moderate Strong Rudolph 2008 Benztropine (oral) prevention of Trihexyphenidyl extrapyramidal symptoms with antipsychotics; more effective agents available for treatment of Parkinson disease. Antispasmodics Highly anticholinergic, Avoid Moderate Strong Lechevallier- Belladonna alkaloids uncertain except in Michel 2005 Clidinium-chlordiazepoxide effectiveness. short-term Rudolph 2008 Dicyclomine palliative Hyoscyamine care to Propantheline decrease Scopolamine oral secretions. Antithrombotics Dipyridamole, oral short-acting* May -

Characterisation of the Α1b-Adrenoceptor by Modeling, Dynamics and Virtual Screening Kapil Jain B.Pharm, M.S.(Pharm.)
Characterisation of the α1B-Adrenoceptor by Modeling, Dynamics and Virtual Screening Kapil Jain B.Pharm, M.S.(Pharm.) A Thesis submitted for the degree of Master of Philosophy at The University of Queensland in 2018 Institute for Molecular Bioscience 0 Abstract G protein-coupled receptors (GPCRs) are the largest druggable class of proteins yet relatively little is known about the mechanism by which agonist binding induces the conformational changes necessary for G protein activation and intracellular signaling. Recently, the Kobilka group has shown that agonists, neutral antagonists and inverse agonists stabilise distinct extracellular surface (ECS) conformations of the β2-adrenergic receptor (AR) opening up new possibilities for allosteric drug targeting at GPCRs. The goal of this project is to extend these studies to define how the ECS conformation of the α1B-AR changes during agonist binding and develop an understanding of ligand entry and exit mechanisms that may help in the design of specific ligands with higher selectivity, efficacy and longer duration of action. Two parallel approaches were initiated to identify likely functional residues. The role of residues lining the primary binding site were predicted by online web server (Q-Site Finder) while secondary binding sites residues were predicted from molecular dynamics (MD) simulations. Predicted functionally significant residues were mutated and their function was established using FLIPR, radioligand and saturation binding assays. Despite the α1B-AR being pursued as a drug target for over last few decades, few specific agonists and antagonists are known to date. In an attempt to address this gap, we pursued ligand-based approach to find potential new leads. -

(12) United States Patent (10) Patent No.: US 9,498,481 B2 Rao Et Al
USOO9498481 B2 (12) United States Patent (10) Patent No.: US 9,498,481 B2 Rao et al. (45) Date of Patent: *Nov. 22, 2016 (54) CYCLOPROPYL MODULATORS OF P2Y12 WO WO95/26325 10, 1995 RECEPTOR WO WO99/O5142 2, 1999 WO WOOO/34283 6, 2000 WO WO O1/92262 12/2001 (71) Applicant: Apharaceuticals. Inc., La WO WO O1/922.63 12/2001 olla, CA (US) WO WO 2011/O17108 2, 2011 (72) Inventors: Tadimeti Rao, San Diego, CA (US); Chengzhi Zhang, San Diego, CA (US) OTHER PUBLICATIONS Drugs of the Future 32(10), 845-853 (2007).* (73) Assignee: Auspex Pharmaceuticals, Inc., LaJolla, Tantry et al. in Expert Opin. Invest. Drugs (2007) 16(2):225-229.* CA (US) Wallentin et al. in the New England Journal of Medicine, 361 (11), 1045-1057 (2009).* (*) Notice: Subject to any disclaimer, the term of this Husted et al. in The European Heart Journal 27, 1038-1047 (2006).* patent is extended or adjusted under 35 Auspex in www.businesswire.com/news/home/20081023005201/ U.S.C. 154(b) by Od en/Auspex-Pharmaceuticals-Announces-Positive-Results-Clinical M YW- (b) by ayS. Study (published: Oct. 23, 2008).* This patent is Subject to a terminal dis- Concert In www.concertpharma. com/news/ claimer ConcertPresentsPreclinicalResultsNAMS.htm (published: Sep. 25. 2008).* Concert2 in Expert Rev. Anti Infect. Ther. 6(6), 782 (2008).* (21) Appl. No.: 14/977,056 Springthorpe et al. in Bioorganic & Medicinal Chemistry Letters 17. 6013-6018 (2007).* (22) Filed: Dec. 21, 2015 Leis et al. in Current Organic Chemistry 2, 131-144 (1998).* Angiolillo et al., Pharmacology of emerging novel platelet inhibi (65) Prior Publication Data tors, American Heart Journal, 2008, 156(2) Supp. -

Appendix A: Potentially Inappropriate Prescriptions (Pips) for Older People (Modified from ‘STOPP/START 2’ O’Mahony Et Al 2014)
Appendix A: Potentially Inappropriate Prescriptions (PIPs) for older people (modified from ‘STOPP/START 2’ O’Mahony et al 2014) Consider holding (or deprescribing - consult with patient): 1. Any drug prescribed without an evidence-based clinical indication 2. Any drug prescribed beyond the recommended duration, where well-defined 3. Any duplicate drug class (optimise monotherapy) Avoid hazardous combinations e.g.: 1. The Triple Whammy: NSAID + ACE/ARB + diuretic in all ≥ 65 year olds (NHS Scotland 2015) 2. Sick Day Rules drugs: Metformin or ACEi/ARB or a diuretic or NSAID in ≥ 65 year olds presenting with dehydration and/or acute kidney injury (AKI) (NHS Scotland 2015) 3. Anticholinergic Burden (ACB): Any additional medicine with anticholinergic properties when already on an Anticholinergic/antimuscarinic (listed overleaf) in > 65 year olds (risk of falls, increased anticholinergic toxicity: confusion, agitation, acute glaucoma, urinary retention, constipation). The following are known to contribute to the ACB: Amantadine Antidepressants, tricyclic: Amitriptyline, Clomipramine, Dosulepin, Doxepin, Imipramine, Nortriptyline, Trimipramine and SSRIs: Fluoxetine, Paroxetine Antihistamines, first generation (sedating): Clemastine, Chlorphenamine, Cyproheptadine, Diphenhydramine/-hydrinate, Hydroxyzine, Promethazine; also Cetirizine, Loratidine Antipsychotics: especially Clozapine, Fluphenazine, Haloperidol, Olanzepine, and phenothiazines e.g. Prochlorperazine, Trifluoperazine Baclofen Carbamazepine Disopyramide Loperamide Oxcarbazepine Pethidine -

Commonly Prescribed Psychotropic Medications
COMMONLY PRESCRIBED PSYCHOTROPIC MEDICATIONS NAME Generic (Trade) DOSAGE KEY CLINICAL INFORMATION Antidepressant Medications* Start: IR-100 mg bid X 4d then ↑ to 100 mg tid; SR-150 mg qam X 4d then ↑ to 150 mg Contraindicated in seizure disorder because it decreases seizure threshold; stimulating; not good for treating anxiety disorders; second Bupropion (Wellbutrin) bid; XL-150 mg qam X 4d, then ↑ to 300 mg qam. Range: 300-450 mg/d. line TX for ADHD; abuse potential. ¢ (IR/SR), $ (XL) Citalopram (Celexa) Start: 10-20 mg qday,↑10-20 mg q4-7d to 30-40 mg qday. Range: 20-60 mg/d. Best tolerated of SSRIs; very few and limited CYP 450 interactions; good choice for anxious pt. ¢ Duloxetine (Cymbalta) Start: 30 mg qday X 1 wk, then ↑ to 60 mg qday. Range: 60-120 mg/d. More GI side effects than SSRIs; tx neuropathic pain; need to monitor BP; 2nd line tx for ADHD. $ Escitalopram (Lexapro) Start: 5 mg qday X 4-7d then ↑ to 10 mg qday. Range 10-30 mg/d (3X potent vs. Celexa). Best tolerated of SSRIs, very few and limited CYP 450 interactions. Good choice for anxious pt. $ Fluoxetine (Prozac) Start: 10 mg qam X 4-7d then ↑ to 20 mg qday. Range: 20-60 mg/d. More activating than other SSRIs; long half-life reduces withdrawal (t ½ = 4-6 d). ¢ Mirtazapine (Remeron) Start: 15 mg qhs. X 4-7d then ↑ to 30 mg qhs. Range: 30-60 mg/qhs. Sedating and appetite promoting; Neutropenia risk (1 in 1000) so avoid in immunosupressed patients. -

(19) United States (12) Patent Application Publication (10) Pub
US 20130289061A1 (19) United States (12) Patent Application Publication (10) Pub. No.: US 2013/0289061 A1 Bhide et al. (43) Pub. Date: Oct. 31, 2013 (54) METHODS AND COMPOSITIONS TO Publication Classi?cation PREVENT ADDICTION (51) Int. Cl. (71) Applicant: The General Hospital Corporation, A61K 31/485 (2006-01) Boston’ MA (Us) A61K 31/4458 (2006.01) (52) U.S. Cl. (72) Inventors: Pradeep G. Bhide; Peabody, MA (US); CPC """"" " A61K31/485 (201301); ‘4161223011? Jmm‘“ Zhu’ Ansm’ MA. (Us); USPC ......... .. 514/282; 514/317; 514/654; 514/618; Thomas J. Spencer; Carhsle; MA (US); 514/279 Joseph Biederman; Brookline; MA (Us) (57) ABSTRACT Disclosed herein is a method of reducing or preventing the development of aversion to a CNS stimulant in a subject (21) App1_ NO_; 13/924,815 comprising; administering a therapeutic amount of the neu rological stimulant and administering an antagonist of the kappa opioid receptor; to thereby reduce or prevent the devel - . opment of aversion to the CNS stimulant in the subject. Also (22) Flled' Jun‘ 24’ 2013 disclosed is a method of reducing or preventing the develop ment of addiction to a CNS stimulant in a subj ect; comprising; _ _ administering the CNS stimulant and administering a mu Related U‘s‘ Apphcatlon Data opioid receptor antagonist to thereby reduce or prevent the (63) Continuation of application NO 13/389,959, ?led on development of addiction to the CNS stimulant in the subject. Apt 27’ 2012’ ?led as application NO_ PCT/US2010/ Also disclosed are pharmaceutical compositions comprising 045486 on Aug' 13 2010' a central nervous system stimulant and an opioid receptor ’ antagonist. -

M100907, a Serotonin 5-HT2A Receptor Antagonist and Putative Antipsychotic, Blocks Dizocilpine-Induced Prepulse Inhibition Defic
M100907, a Serotonin 5-HT2A Receptor Antagonist and Putative Antipsychotic, Blocks Dizocilpine-Induced Prepulse Inhibition Deficits in Sprague–Dawley and Wistar Rats Geoffrey B. Varty, Ph.D., Vaishali P. Bakshi, Ph.D., and Mark A. Geyer, Ph.D. a In a recent study using Wistar rats, the serotonergic 5-HT2 1 receptor agonist cirazoline disrupts PPI. As risperidone a receptor antagonists ketanserin and risperidone reduced the and M100907 have affinity at the 1 receptor, a final study disruptive effects of the noncompetitive N-methyl-D- examined whether M100907 would block the effects of aspartate (NMDA) antagonist dizocilpine on prepulse cirazoline on PPI. Risperidone partially, but inhibition (PPI), suggesting that there is an interaction nonsignificantly, reduced the effects of dizocilpine in Wistar between serotonin and glutamate in the modulation of PPI. rats, although this effect was smaller than previously In contrast, studies using the noncompetitive NMDA reported. Consistent with previous studies, risperidone did antagonist phencyclidine (PCP) in Sprague–Dawley rats not alter the effects of dizocilpine in Sprague–Dawley rats. found no effect with 5-HT2 antagonists. To test the hypothesis Most importantly, M100907 pretreatment fully blocked the that strain differences might explain the discrepancy in effect of dizocilpine in both strains; whereas SDZ SER 082 these findings, risperidone was tested for its ability to had no effect. M100907 had no influence on PPI by itself reduce the PPI-disruptive effects of dizocilpine in Wistar and did not reduce the effects of cirazoline on PPI. These and Sprague–Dawley rats. Furthermore, to determine studies confirm the suggestion that serotonin and glutamate which serotonergic receptor subtype may mediate this effect, interact in modulating PPI and indicate that the 5-HT2A the 5-HT2A receptor antagonist M100907 (formerly MDL receptor subtype mediates this interaction. -
![Selective Labeling of Serotonin Receptors Byd-[3H]Lysergic Acid](https://docslib.b-cdn.net/cover/9764/selective-labeling-of-serotonin-receptors-byd-3h-lysergic-acid-319764.webp)
Selective Labeling of Serotonin Receptors Byd-[3H]Lysergic Acid
Proc. Nati. Acad. Sci. USA Vol. 75, No. 12, pp. 5783-5787, December 1978 Biochemistry Selective labeling of serotonin receptors by d-[3H]lysergic acid diethylamide in calf caudate (ergots/hallucinogens/tryptamines/norepinephrine/dopamine) PATRICIA M. WHITAKER AND PHILIP SEEMAN* Department of Pharmacology, University of Toronto, Toronto, Canada M5S 1A8 Communicated by Philip Siekevltz, August 18,1978 ABSTRACT Since it was known that d-lysergic acid di- The objective in this present study was to improve the se- ethylamide (LSD) affected catecholaminergic as well as sero- lectivity of [3H]LSD for serotonin receptors, concomitantly toninergic neurons, the objective in this study was to enhance using other drugs to block a-adrenergic and dopamine receptors the selectivity of [3HJISD binding to serotonin receptors in vitro by using crude homogenates of calf caudate. In the presence of (cf. refs. 36-38). We then compared the potencies of various a combination of 50 nM each of phentolamine (adde to pre- drugs on this selective [3H]LSD binding and compared these clude the binding of [3HJLSD to a-adrenoceptors), apmo ie, data to those for the high-affinity binding of [3H]serotonin and spiperone (added to preclude the binding of [3H[LSD to (39). dopamine receptors), it was found by Scatchard analysis that the total number of 3H sites went down to 300 fmol/mg, compared to 1100 fmol/mg in the absence of the catechol- METHODS amine-blocking drugs. The IC50 values (concentrations to inhibit Preparation of Membranes. Calf brains were obtained fresh binding by 50%) for various drugs were tested on the binding of [3HLSD in the presence of 50 nM each of apomorphine (A), from the Canada Packers Hunisett plant (Toronto). -

United States Patent (19) (11) 4,310,524 Wiech Et Al
United States Patent (19) (11) 4,310,524 Wiech et al. 45 Jan. 12, 1982 (54) TCA COMPOSITION AND METHOD FOR McMillen et al., Fed. Proc., 38,592 (1979). RAPD ONSET ANTDEPRESSANT Sellinger et al., Fed. Proc., 38,592 (1979). THERAPY Pandey et al., Fed. Proc., 38,592 (1979). 75) Inventors: Norbert L. Wiech; Richard C. Ursillo, Primary Examiner-Stanley J. Friedman both of Cincinnati, Ohio Attorney, Agent, or Firm-Millen & White 73) Assignee: Richardson-Merrell, Inc., Wilton, Conn. (57 ABSTRACT A method is provided for treating depression in a pa (21) Appl. No.: 139,498 tient therefrom and requiring rapid symptomatic relief, (22 Filed: Apr. 11, 1980 which comprises administering to said patient concur 51) Int. Cl. .................... A61K 31/33; A61K 31/135 rently (a) an effective antidepressant amount of a tricy clic antidepressant or a pharmaceutically effective acid (52) ...... 424/244; 424/330 addition salt thereof, and (b) an amount of an a-adrener 58) Field of Search ................................ 424/244, 330 gic receptor blocking agent effective to achieve rapid (56) References Cited onset of the antidepressant action of (a), whereby the PUBLICATIONS onset of said antidepressant action is achieved within Chemical Abst., vol. 66-72828m, (1967), Kellett. from 1 to 7 days. Chemical Abst, vol. 68-94371a, (1968), Martelli et al. A pharmaceutical composition is also provided which is Chemical Abst., vol. 74-86.048j, (1971), Dixit et al. especially adapted for use with the foregoing method. Holmberg et al., Psychopharm., 2,93 (1961). Svensson, Symp. Med. Hoechst., 13, 245 (1978). 17 Claims, No Drawings 4,310,524 1. -

4 Supplementary File
Supplemental Material for High-throughput screening discovers anti-fibrotic properties of Haloperidol by hindering myofibroblast activation Michael Rehman1, Simone Vodret1, Luca Braga2, Corrado Guarnaccia3, Fulvio Celsi4, Giulia Rossetti5, Valentina Martinelli2, Tiziana Battini1, Carlin Long2, Kristina Vukusic1, Tea Kocijan1, Chiara Collesi2,6, Nadja Ring1, Natasa Skoko3, Mauro Giacca2,6, Giannino Del Sal7,8, Marco Confalonieri6, Marcello Raspa9, Alessandro Marcello10, Michael P. Myers11, Sergio Crovella3, Paolo Carloni5, Serena Zacchigna1,6 1Cardiovascular Biology, 2Molecular Medicine, 3Biotechnology Development, 10Molecular Virology, and 11Protein Networks Laboratories, International Centre for Genetic Engineering and Biotechnology (ICGEB), Padriciano, 34149, Trieste, Italy 4Institute for Maternal and Child Health, IRCCS "Burlo Garofolo", Trieste, Italy 5Computational Biomedicine Section, Institute of Advanced Simulation IAS-5 and Institute of Neuroscience and Medicine INM-9, Forschungszentrum Jülich GmbH, 52425, Jülich, Germany 6Department of Medical, Surgical and Health Sciences, University of Trieste, 34149 Trieste, Italy 7National Laboratory CIB, Area Science Park Padriciano, Trieste, 34149, Italy 8Department of Life Sciences, University of Trieste, Trieste, 34127, Italy 9Consiglio Nazionale delle Ricerche (IBCN), CNR-Campus International Development (EMMA- INFRAFRONTIER-IMPC), Rome, Italy This PDF file includes: Supplementary Methods Supplementary References Supplementary Figures with legends 1 – 18 Supplementary Tables with legends 1 – 5 Supplementary Movie legends 1, 2 Supplementary Methods Cell culture Primary murine fibroblasts were isolated from skin, lung, kidney and hearts of adult CD1, C57BL/6 or aSMA-RFP/COLL-EGFP mice (1) by mechanical and enzymatic tissue digestion. Briefly, tissue was chopped in small chunks that were digested using a mixture of enzymes (Miltenyi Biotec, 130- 098-305) for 1 hour at 37°C with mechanical dissociation followed by filtration through a 70 µm cell strainer and centrifugation. -

Prazosin: Preliminary Report and Comparative Studies with Other
298 BRITISH MEDICAL JOURNAL 1 1 MAY 1974 Prazosin: Preliminary Report and Comparative Studies with Other Antihypertensive Agents Br Med J: first published as 10.1136/bmj.2.5914.298 on 11 May 1974. Downloaded from GORDON S. STOKES, MICHAEL A. WEBER British Medical Journal, 1974, 2, 298-300 bilirubin. One patient had minimal grade 3 hypertensive ocu- lar fundus changes, 10 had grade 1 or 2 changes, and four had normal fundi. Serum crea-tinine concentration was normal in Summary every patient. A diagnosis of uncomplicated essential hypertension was In a group of 14 hypertensive patients a 10-week course made in 12 patients. In two other patients pyelographic of treatment with prazosin 3-7 5 mg/day produced a signifi- evidence of analgesic nephropathy was found. The remaining cant reduction in mean blood pressure without serious side patient had essential hypertension and stable angina pectoris. effects. The fall in diastolic pressure exceeded the response After at least two baseline blood pressure readings, taken mm more to a placebo by 10 Hg or in 9 patients. The average when patients were recumbent and standing, treatment was decrease in diastolic pressure was similar to that produced started with prazosin, 1-mg capsules by mouth, three times or by methyldopa 750 mg/day propranolol 120-160 mg/day daily. Thereafter the patient visited the clinic at intervals of was but the fall in systolic pressure comparatively smaller, one to three weeks. At each visit blood pressure was meas- that the consistent with reported experimental work showing ured to the nearest 5 mm Hg with an Accoson sphygmomo- drug causes vasodilatation. -

Ring-Substituted Amphetamine Interactions with Neurotransmitter Receptor Binding Sites in Human Cortex
208 NeuroscienceLetters, 95 (1988) 208-212 Elsevier Scientific Publishers Ireland Ltd. NSL O5736 Ring-substituted amphetamine interactions with neurotransmitter receptor binding sites in human cortex Pamela A. Pierce and Stephen J. Peroutka Deparmentsof Neurologyand Pharmacology, Stanford University Medical Center, StanJbrd,CA 94305 (U.S.A.) (Received 31 May 1988; Revised version received 11 August 1988; Accepted 12 August 1988) Key words.' Ring-substituted amphetamine; (_+)-3,4-Methylenedioxyamphetamine; ( +_)-3,4-Methylene- dioxyethamphetamine; (_+)-3,4-Methylenedioxymethamphetamine; Ecstasy; 4-Bromo-2,5- dimethoxyphenylisopropylamine binding site; Human cortical receptor The binding affinities of 3 ring-substituted amphetamine compounds were determined at 9 neurotrans- mitter binding sites in human cortex. (_+)-3,4-Methylenedioxyamphetamine (MDAk (_+)-3,4-methylene- dioxyethamphetamine (MDE), and (_+)-3,4-methylenedioxymethamphetamine (MDMA or 'Ecstasy') all display highest affinity (approximately 1 /tM) for the recently identified 'DOB binding site' labeled by ['TBr]R(-)4-bromo-2,5-dimethoxyphenylisopropylamine ([77Br]R(-)DOB). MDA displays moderate affinity (4-5/iM) for the 5-hydroxytryptaminetA (5-HTr^), 5-HT_D,and =,-adrenergic sites in human cor- tex. MDE and MDMA display lower affinity or are inactive at all other sites tested in the present study. These observations are discussed in relation to the novel psychoactive effects of the ring-substituted amphetamines. A series of ring-substituted amphetamine derivatives exist which are structurally related to both amphetamines and hallucinogens. However, drugs such as (+)-3,4- methylenedioxyamphetamine (MDA), (_+)-3,4-methylenedioxyethamphetamine (MDE), and (+)-3,4-methylenedioxymethamphetamine (MDMA or 'Ecstasy') ap- pear to produce unique psychoactive effects which are distinct from the effects of both amphetamines and hallucinogens [13, 15].