Comparative Analysis of Actaea Chloroplast Genomes and Molecular Marker Development for the Identification of Authentic Cimicifugae Rhizoma
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Extended Glossary 11 Extended Glossary
Extended Glossary 11 EXTENDED GLOSSARY This glossary combines some of the helpful found on the RHS website at www.rhs.org.uk/ introductory sections from older editions in an agmplants. alphabetical listing. A fuller, more discursive account The AGM plant list has, to date, been re-examined of plant names, Guide to Plant Names, and a detailed every 10 years. The latest review was carried out guide to the typography of plant names, during 2012 and published in February 2013. Recommended Style for Printing Plant Names, are both available as RHS Advisory Leaflets. To request Botanical Names a copy of either please send an A4 sae to The Compiler at the contact address given on page 5. The aim of the botanical naming system is to provide each different plant with a single, unique, universal Advisory Committee on name. The basic unit of plant classification is the Nomenclature and Taxonomy species. Species that share a number of significant characteristics are grouped together to form a genus See under the new name of Nomenclature and (plural genera). The name of a species is made up of Taxonomy Advisory Group two elements; the name of the genus followed by the specific epithet, for example, Narcissus romieuxii. Authorities Variation within a species can be recognised by division into subspecies (usually abbreviated to In order that plant names can be used with precision subsp.), varietas (or variety abbreviated to var.) and throughout the scientific world, the name of the forma (or form abbreviated to f.). Whilst it is person who coined the name of a plant species (its unusual for a plant to have all of these, it is possible, author, or authority) is added to the plant name. -
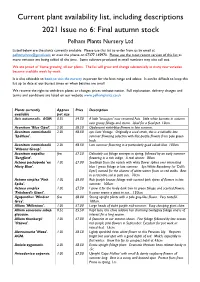
Current Plant Availability List, Including Descriptions 2021 Issue No 6: Final Autumn Stock Pelham Plants Nursery Ltd
Current plant availability list, including descriptions 2021 Issue no 6: Final autumn stock Pelham Plants Nursery Ltd Listed below are the plants currently available. Please use this list to order from us by email at [email protected] or over the phone on 07377 145970. Please use the most recent version of this list as more varieties are being added all the time. Some cultivars produced in small numbers may also sell out. We are proud of ‘home growing’ all our plants. The list will grow and change substantially as many new varieties become available week by week. It is also advisable to book to visit the nursery in person for the best range and advice. It can be difficult to keep this list up to date at our busiest times or when batches are small. We reserve the right to withdraw plants or changes prices without notice. Full explanation, delivery charges and terms and conditions are listed on our website www.pelhamplants.co.uk Plants currently Approx Price Description available pot size Acis autumnalis. AGM. 0.5L £4.50 A little 'Leucojum' now renamed Acis. Little white bonnets in autumn over grassy foliage and stems. Ideal for a focal pot. 10cm. Aconitum 'Blue Opal'. 2.0L £8.50 Opalescent violet-blue flowers in late summer. Aconitum carmichaelii 2.0L £8.50 syn. Late Vintage. Originally a seed strain, this is a valuable late 'Spätlese'. summer flowering selection with lilac-purple flowers from pale green buds. Aconitum carmichaelii 2.0L £8.50 Late summer flowering in a particularly good cobalt blue. -

Preparing the Shaanxi-Qinling Mountains Integrated Ecosystem Management Project (Cofinanced by the Global Environment Facility)
Technical Assistance Consultant’s Report Project Number: 39321 June 2008 PRC: Preparing the Shaanxi-Qinling Mountains Integrated Ecosystem Management Project (Cofinanced by the Global Environment Facility) Prepared by: ANZDEC Limited Australia For Shaanxi Province Development and Reform Commission This consultant’s report does not necessarily reflect the views of ADB or the Government concerned, and ADB and the Government cannot be held liable for its contents. (For project preparatory technical assistance: All the views expressed herein may not be incorporated into the proposed project’s design. FINAL REPORT SHAANXI QINLING BIODIVERSITY CONSERVATION AND DEMONSTRATION PROJECT PREPARED FOR Shaanxi Provincial Government And the Asian Development Bank ANZDEC LIMITED September 2007 CURRENCY EQUIVALENTS (as at 1 June 2007) Currency Unit – Chinese Yuan {CNY}1.00 = US $0.1308 $1.00 = CNY 7.64 ABBREVIATIONS ADB – Asian Development Bank BAP – Biodiversity Action Plan (of the PRC Government) CAS – Chinese Academy of Sciences CASS – Chinese Academy of Social Sciences CBD – Convention on Biological Diversity CBRC – China Bank Regulatory Commission CDA - Conservation Demonstration Area CNY – Chinese Yuan CO – company CPF – country programming framework CTF – Conservation Trust Fund EA – Executing Agency EFCAs – Ecosystem Function Conservation Areas EIRR – economic internal rate of return EPB – Environmental Protection Bureau EU – European Union FIRR – financial internal rate of return FDI – Foreign Direct Investment FYP – Five-Year Plan FS – Feasibility -

Lenka Kočková
MASARYKOVA UNIVERZITA PŘÍRODOVĚDECKÁ FAKULTA ÚSTAV BOTANIKY A ZOOLOGIE Velikost genomu a poměr bazí v genomu v čeledi Ranunculaceae Diplomová práce Lenka Kočková Vedoucí práce: Doc. RNDr. Petr Bureš, Ph. D. Brno 2012 Bibliografický záznam Autor: Bc. Lenka Kočková Přírodovědecká fakulta, Masarykova univerzita, Ústav botaniky a zoologie Název práce: Velikost genomu a poměr bazí v genomu v čeledi Ranunculaceae Studijní program: Biologie Studijní obor: Systematická biologie a ekologie (Botanika) Vedoucí práce: Doc. RNDr. Petr Bureš, Ph. D. Akademický rok: 2011/2012 Počet stran: 104 Klíčová slova: Ranunculaceae, průtoková cytometrie, PI/DAPI, DNA obsah, velikost genomu, GC obsah, zastoupení bazí, velikost průduchů, Pignattiho indikační hodnoty Bibliographic Entry Author: Bc. Lenka Kočková Faculty of Science, Masaryk University, Department of Botany and Zoology Title of Thesis: Genome size and genomic base composition in Ranunculaceae Programme: Biology Field of Study: Systematic Biology and Ecology (Botany) Supervisor: Doc. RNDr. Petr Bureš, Ph. D. Academic Year: 2011/2012 Number of Pages: 104 Keywords: Ranunculaceae, flow cytometry, PI/DAPI, DNA content, genome size, GC content, base composition, stomatal size, Pignatti‘s indicator values Abstrakt Pomocí průtokové cytometrie byla změřena velikost genomu a AT/GC genomový poměr u 135 druhů z čeledi Ranunculaceae. U druhů byla naměřena délka a šířka průduchů a z literatury byly získány údaje o počtu chromozomů a ekologii druhů. Velikost genomu se v rámci čeledi liší 63-krát. Nejmenší genom byl naměřen u Aquilegia canadensis (2C = 0,75 pg), největší u Ranunculus lingua (2C = 47,93 pg). Mezi dvěma hlavními podčeleděmi Ranunculoideae a Thalictroideae je ve velikosti genomu markantní rozdíl (2C = 2,48 – 47,94 pg a 0,75 – 4,04 pg). -

Gymnaconitum, a New Genus of Ranunculaceae Endemic to the Qinghai-Tibetan Plateau
TAXON 62 (4) • August 2013: 713–722 Wang & al. • Gymnaconitum, a new genus of Ranunculaceae Gymnaconitum, a new genus of Ranunculaceae endemic to the Qinghai-Tibetan Plateau Wei Wang,1 Yang Liu,2 Sheng-Xiang Yu,1 Tian-Gang Gao1 & Zhi-Duan Chen1 1 State Key Laboratory of Systematic and Evolutionary Botany, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, P.R. China 2 Department of Ecology and Evolutionary Biology, University of Connecticut, Storrs, Connecticut 06269-3043, U.S.A. Author for correspondence: Wei Wang, [email protected] Abstract The monophyly of traditional Aconitum remains unresolved, owing to the controversial systematic position and taxonomic treatment of the monotypic, Qinghai-Tibetan Plateau endemic A. subg. Gymnaconitum. In this study, we analyzed two datasets using maximum likelihood and Bayesian inference methods: (1) two markers (ITS, trnL-F) of 285 Delphinieae species, and (2) six markers (ITS, trnL-F, trnH-psbA, trnK-matK, trnS-trnG, rbcL) of 32 Delphinieae species. All our analyses show that traditional Aconitum is not monophyletic and that subgenus Gymnaconitum and a broadly defined Delphinium form a clade. The SOWH tests also reject the inclusion of subgenus Gymnaconitum in traditional Aconitum. Subgenus Gymnaconitum markedly differs from other species of Aconitum and other genera of tribe Delphinieae in many non-molecular characters. By integrating lines of evidence from molecular phylogeny, divergence times, morphology, and karyology, we raise the mono- typic A. subg. Gymnaconitum to generic status. Keywords Aconitum; Delphinieae; Gymnaconitum; monophyly; phylogeny; Qinghai-Tibetan Plateau; Ranunculaceae; SOWH test Supplementary Material The Electronic Supplement (Figs. S1–S8; Appendices S1, S2) and the alignment files are available in the Supplementary Data section of the online version of this article (http://www.ingentaconnect.com/content/iapt/tax). -

2019 Program WELCOME
THE SCOTT ARBORETUM OF SWARTHMORE COLLEGE www.scottarboretum.org 2019 Program WELCOME Welcome TABLE OF CONTENTS Greetings! Welcome to the 2019 Scott Arboretum Selections: Spring Sale. Download this handbook at scottarboretum.org. WELCOME 2 Schedule of the Sale 3 Special Offer Special Friends 4 10% discount on sales $100 and over, applies to plants only. Planting Container Grown Plants 10 Meaning of our Labels 12 Refund Policy Plant List 13 ALL SALES ARE FINAL; NO EXCHANGES OR REFUNDS. We are not able to offer refunds or exchanges since this is a special once-a- year event. Thank you! Many thanks to those volunteers who have contributed their efforts to this sale. A special thank you to Alan Kruza and Eve Thryum whose unwavering support and passion for the plants makes this sale possible. 2 SCHEDULE OF THE SALE Scott Arboretum Selections: Spring Sale Schedule: Friday, May 10 Special Friends Preview Party 5:30 to 7:30 pm To become a Special Friend to attend our Preview Party, call the Scott Arboretum Offices at 610- 328-8025. Saturday, May 11 Members Shopping 10 am – noon Members must show their membership card for early admission. If you have lost or misplaced your card, or would like to become a member, please call 610-328-8025. Open to the public – free noon – 3 pm 3 SPECIAL FRIENDS Julia and Vincent Auletta Our sincere appreciation to William D. Conwell Charles and Rosemary Philips these Special Friends of the Scott Laura Axel Arboretum Selections Sales, whose Harold Sweetman Alice Reilly support helps underwrite the cost of these vital fund-raising events. -

Literaturverzeichnis
Literaturverzeichnis Abaimov, A.P., 2010: Geographical Distribution and Ackerly, D.D., 2009: Evolution, origin and age of Genetics of Siberian Larch Species. In Osawa, A., line ages in the Californian and Mediterranean flo- Zyryanova, O.A., Matsuura, Y., Kajimoto, T. & ras. Journal of Biogeography 36, 1221–1233. Wein, R.W. (eds.), Permafrost Ecosystems. Sibe- Acocks, J.P.H., 1988: Veld Types of South Africa. 3rd rian Larch Forests. Ecological Studies 209, 41–58. Edition. Botanical Research Institute, Pretoria, Abbadie, L., Gignoux, J., Le Roux, X. & Lepage, M. 146 pp. (eds.), 2006: Lamto. Structure, Functioning, and Adam, P., 1990: Saltmarsh Ecology. Cambridge Uni- Dynamics of a Savanna Ecosystem. Ecological Stu- versity Press. Cambridge, 461 pp. dies 179, 415 pp. Adam, P., 1994: Australian Rainforests. Oxford Bio- Abbott, R.J. & Brochmann, C., 2003: History and geography Series No. 6 (Oxford University Press), evolution of the arctic flora: in the footsteps of Eric 308 pp. Hultén. Molecular Ecology 12, 299–313. Adam, P., 1994: Saltmarsh and mangrove. In Groves, Abbott, R.J. & Comes, H.P., 2004: Evolution in the R.H. (ed.), Australian Vegetation. 2nd Edition. Arctic: a phylogeographic analysis of the circu- Cambridge University Press, Melbourne, pp. marctic plant Saxifraga oppositifolia (Purple Saxi- 395–435. frage). New Phytologist 161, 211–224. Adame, M.F., Neil, D., Wright, S.F. & Lovelock, C.E., Abbott, R.J., Chapman, H.M., Crawford, R.M.M. & 2010: Sedimentation within and among mangrove Forbes, D.G., 1995: Molecular diversity and deri- forests along a gradient of geomorphological set- vations of populations of Silene acaulis and Saxi- tings. -

Botanical Name Common Name Type Description Actaea Simplex 'Hillside Black Beauty' Bugbane Perennial Grow in Part Shade to Shade
Botanical Name Common Name Type Description Actaea simplex 'Hillside Black Beauty' Bugbane Perennial Grow in part shade to shade. Best in groups. Creamy white frangant flowers. Alcea 'Blacknight' Hollyhock Perennial TrueFull sun perennial Hollyhock! Alcea 'Las Vegas' Hollyhock Biennial Mix of colorful flowers like Vegas! Alcea 'Mars Magic' Hollyhock Perennial True full sun perennial Hollyhock! Allium 'Millenium' Ornamental Onion Perennial 2018 Plant of the Year! Rosy Purple 2" ball Blooms. Anemone 'Whirlwind' Japanese Anemone Perennial Very large showy snow-white poppy blossoms. Aquilegia canadensis Columbine Perennial One of the frist flowers to provide nectar! Asclepias tuberosa Butterfly Weed Perennial Place this in a sunny spot for a pop of bright orange to attact butterflies. Astilbe 'Amber Moon' False Spirea Perennial Rasberry pink plumes with gold foliageand lacy texture. Astilbe 'Snowdrift' False Spirea Perennial White showy blooms with fern-like foliage! Astilbe 'Vision in Red' False Spirea Perennial Fuzzy deep red dense plumes with red stems. Athyrium 'Ghost' Lady Fern Fern Slivery upright foliage that does best in light shade. Giant Japanese Painted Athyrium 'Godzilla' Fern Fern Monster growing painted fern Athyrium filix-femina var. angustum Lacy light green foliage with upright dark red stems. 'Lady in Red' Northern Lady Fern Fern Outstanding painted showpiece fern! Athyrium niponicum 'Regal Red' Japanese Painted Fern Fern Baptisia 'Dutch Chocolate' Decadence® Series False Indigo Perennial Easy to grow with ornamental seed pods. Baptisia 'Lemon Meringue' Decadence® Series False Indigo Perennial Gorgeous long lasting lemon yellow flowers. Caryopteris x clandonensis 'Blue Empire' Bluebeard Shrub Bigger blue flower clusters Caryopteris x clandonensis 'Dark Knight' Bluebeard Shrub Showy,fragrant, and easy to grow. -

Garden Wise Non-Invasive Plants for Your Garden
Garden Wise Non-Invasive Plants for Your Garden Western Washington Guide Voluntary codes of conduct For the gardening public (annotated): In an effort to reduce the spread of invasive plants used for horticultural purposes, experts have created the “Voluntary Codes of Conduct,” a series of steps that nursery professionals, landscape architects, gardeners, and others can take to help curb the spread of invasive horticultural plants. ◊ Ask for only non-invasive species when you acquire plants. Plant only environmentally safe species in your gardens. Work towards and promote new landscape design that is friendly to regional ecosystems. ◊ Seek information on which species are invasive in your area. Sources could include botanical gardens, horticulturists, conservationists, and government agencies. Remove invasive species from your land and replace them with non-invasive species suited to your site and needs. ◊ Do not trade plants with other gardeners if you know they are species with invasive characteristics. ◊ Request that botanical gardens and nurseries promote, display, and sell only non-invasive species. ◊ Help educate your community and other gardeners in your area through personal contact and in such settings as garden clubs and other civic groups. For the full Gardening Codes of Conduct, or to learn about the Codes of Conduct for Government, Nursery Professionals, Landscape Architects, and Botanic Gardens and Arboreta, please go to the Invasive.org, TNC’s Global Invasive Species Team webpage: www.invasive.org/gist/horticulture/using-codes.html. Garden Wise is dedicated to Ann Lennartz Garden Wise Non-Invasive Plants for Your Garden While most exotic plants are not problematic, a few have become invasive in Washington State. -

112 – April 2009 Newsletter
The Irish Garden Plant Society Newsletter No. 112 April 2009 In This Issue 1 Editorial 2 Letter from the Chairman 3 The Lismacloskey Rectory Garden & Project Irish cultivar conservation by Patrick Quigley 6 A Dangerous Walk with Bob Bradshaw 8 John Joe Costin introduces Broadleaved Evergreen Trees 15 Rae McIntyre Reminiscing 19 Worth a Read by Paddy Tobin 24 Collectors’ Corner Bulbinella hookeri Peter Milligan & Nicola Milligan 28 Details of the Annual General Meeting 31 Gail Roantree visits the 2008 Gothenburg International Garden Festival 33 Seed Exchange Report 2009 by Stephen Butler 34 Seamus O’Briens tells the story of Lilium henryi now 120 years in cultivation 37 Regional Reports 45 Looking Ahead 48 Mary Bradshaw extols ‘Ireland’s Wild Orchids a field guide’ Front cover: Moji Shan known to Augustine Henry and E.H. Wilson as “the Dome”. Henry collected Lilium henryi on its slopes during the 1880s. Lilium henryi in Glasnevin’s Double Herbaceous Borders. Séamus O’Brien Editorial Thank you to everyone who wrote or e-mailed with good wishes over the last few months. The Annual General Meeting takes place next month May 23 rd in Greenmount College Antrim. This is an important forum to discuss the future direction and work of the Society. A new Chairman will be elected as Petronilla Martin’s term of office comes to a close after a busy three years. There are also two vacancies on the National Committee as both Marco Fussy and Carsten Asherfeld have returned to Germany. Their expertise as a garden designer and landscape architect respectively contributed in many ways to the IGPS since they joined the Committee in 2006. -

Master Plant List
MASTER PLANT LIST 5 7 8 6 Glasshouse 4 1 2 3 7 MASTER PLANT LIST PAGE 1 TREES 4 PAPERBARK MAPLE Acer griseum 2 3 RED WEEPING CUT-LEAF JAPANESE MAPLE Acer palmatum ‘Atropurpureum Dissectum’ 3 4 5 7 8 CORAL BARK JAPANESE MAPLE Acer palmatum ‘Sango Kaku’ 4 WEEPING CUT-LEAF JAPANESE MAPLE Acer palmatum ‘Viridis Dissectum’ 2 FULL MOON MAPLE Acer shirasawanum ‘Aureum’ 6 CELESTIAL DOGWOOD Cornus rutgersensis ‘Celestial’ 2 6 SANOMA DOVE TREE Davidia involucrata ‘Sonoma’ 4 SHAKEMASTER HONEY LOCUST Gleditsia triacanthos inermis ‘Shademaster’ 7 TEDDY BEAR MAGNOLIA Magnolia grandiflora ‘Teddy Bear’ 7 BRAKENS BROWN BEAUTY MAGNOLIA Magnolia grandiflora ‘Brackens Brown Beauty’ 2 JAPANESE STEWARTIA Stewartia pseudocamellia 7 WESTERN RED CEDAR Thuja plicata ‘Atrovirens’ SHRUBS 2 ROSANNIE JAPONICA ‘ROZANNIE’ Aucuba japonica ‘Rozannie’ 7 BARBERRY Berberis ‘William Penn’ 2 BEAUTY BERRY Callicarpa ‘Profusion’ 5 7 YULETIDE CAMELLIA Camellia sasanqua ‘Yuletide’ 5 QUINCE Chaenomeles ‘Dragon’s Blood’ 5 QUINCE Chaenomeles ‘Scarlet Storm’ 5 TWIG DOGWOOD WINTER FLAME DOGWOOD Cornus sanguinea ‘Arctic Fire’ 5 MIDWINTER FLAME DOGWOOD Cornus sericea ‘Midwinter Flame’ 1 HARRY LAUDER’S WALKING STICK Corylus avellana ‘Contorta’ 8 BEARBERRY Cotoneaster dammeri 7 SUMMER ICE CAUCASIAN DAPHNE Daphne caucasica ‘Summer Ice’ 2 LILAC DAPHNE Daphne genkwa 6 WINTER DAPHNE Daphne odora f. alba 3 4 CHINESE QUININE Dichroa febrifuga 2 RICE PAPER SHRUB Edgeworthia chrysantha 2 RICE PAPER SHRUB Edgeworhia chrysantha ‘Snow Cream’ 7 TREE IVY Fatshedera lizei 5 DWARF WITCH ALDER Fothergilla gardenii 5 JAPANESE WITCH HAZEL Hamamelis japonica ‘Shibamichi Red’ 2 4 6 BLUE BIRD HYDRANGEA Hydrangea macrophylla ssp. Serrata ‘Bluebird’ 3 4 BLUE DECKLE HYDRANGEA Hydrangea macrophylla ssp. -

Disruption of the Petal Identity Gene APETALA3-3 Is Highly Correlated with Loss of Petals Within the Buttercup Family (Ranunculaceae)
Disruption of the petal identity gene APETALA3-3 is highly correlated with loss of petals within the buttercup family (Ranunculaceae) Rui Zhanga,b,1, Chunce Guoa,b,1, Wengen Zhanga,b,1, Peipei Wanga,b, Lin Lia,b, Xiaoshan Duana,b,c, Qinggao Duc, Liang Zhaoc, Hongyan Shana, Scott A. Hodgesd, Elena M. Kramere, Yi Renc,2, and Hongzhi Konga,2 aState Key Laboratory of Systematic and Evolutionary Botany, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China; bUniversity of Chinese Academy of Sciences, Beijing 100049, China; cCollege of Life Sciences, Shaanxi Normal University, Xi’an 710062, China; dDepartment of Ecology, Evolution and Marine Biology, University of California, Santa Barbara, CA 93106; and eDepartment of Organismic and Evolutionary Biology, Harvard University, Cambridge, MA 02138 Edited by Masatoshi Nei, Pennsylvania State University, University Park, PA, and approved February 11, 2013 (received for review November 12, 2012) Absence of petals, or being apetalous, is usually one of the most plants (including the basal-most lineage, Amborella) (13-16), important features that characterizes a group of flowering plants raises the possibility that the genetic program for petal de- at high taxonomic ranks (i.e., family and above). The apetalous velopment was established before the diversification of extant condition, however, appears to be the result of parallel or angiosperms. Then, during evolution, this program has been fi convergent evolution with unknown genetic causes. Here we modi ed in different directions and to varying degrees, giving show that within the buttercup family (Ranunculaceae), apetalous rise to the production of a huge variety of petals and petaloid genera in at least seven different lineages were all derived from structures (6, 17).