Adrenal Medulla R
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
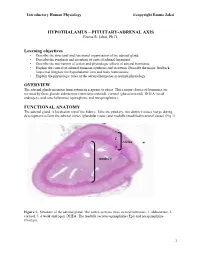
HYPOTHALAMUS – PITUITARY-ADRENAL AXIS Learning Objectives OVERVIEW FUNCTIONAL ANATOMY
Introductory Human Physiology ©copyright Emma Jakoi HYPOTHALAMUS – PITUITARY-ADRENAL AXIS Emma R. Jakoi, Ph.D. Learning objectives • Describe the structural and functional organization of the adrenal gland. • Describe the synthesis and secretion of cortical adrenal hormones. • Describe the mechanism of action and physiologic effects of adrenal hormones. • Explain the control of adrenal hormone synthesis and secretion. Describe the major feedback loops that integrate the hypothalamic axis and body homeostasis. • Explain the physiologic roles of the adrenal hormones in normal physiology. OVERVIEW The adrenal glands maintain homeostasis in response to stress. Three major classes of hormones are secreted by these glands: aldosterone (mineralocorticoid), cortisol (glucocorticoid), DHEA (weak androgen), and catecholamines (epinephrine and norepinephrine). FUNCTIONAL ANATOMY The adrenal gland is located on top of the kidney. Like the pituitary, two distinct tissues merge during development to form the adrenal cortex (glandular tissue) and medulla (modified neuronal tissue) (Fig 1). 1 2 cortex 3 medulla Figure 1. Structure of the adrenal gland. The cortex secretes three steroid hormones: 1. aldosterone, 2. cortisol, 3. a weak androgen, DHEA. The medulla secretes epinephrine (Epi) and norepinephrine (NorEpi). 1 Introductory Human Physiology ©copyright Emma Jakoi MINERALOCORTICOIDS The major mineralocorticoid in humans is aldosterone. Aldosterone is NOT under the hypothalamus- pituitary control and does not mediate a negative feedback to this axis. Aldosterone secretion is increased by the vasoconstrictor, angiotensin II, and by elevated plasma K+ concentration. Elevated plasma Na+ inhibits the secretion of aldosterone. Aldosterone, acts in the kidney to promote secretion of K+ into the urine from the blood and the reabsorption of Na+ from the urine into the blood. -

Vocabulario De Morfoloxía, Anatomía E Citoloxía Veterinaria
Vocabulario de Morfoloxía, anatomía e citoloxía veterinaria (galego-español-inglés) Servizo de Normalización Lingüística Universidade de Santiago de Compostela COLECCIÓN VOCABULARIOS TEMÁTICOS N.º 4 SERVIZO DE NORMALIZACIÓN LINGÜÍSTICA Vocabulario de Morfoloxía, anatomía e citoloxía veterinaria (galego-español-inglés) 2008 UNIVERSIDADE DE SANTIAGO DE COMPOSTELA VOCABULARIO de morfoloxía, anatomía e citoloxía veterinaria : (galego-español- inglés) / coordinador Xusto A. Rodríguez Río, Servizo de Normalización Lingüística ; autores Matilde Lombardero Fernández ... [et al.]. – Santiago de Compostela : Universidade de Santiago de Compostela, Servizo de Publicacións e Intercambio Científico, 2008. – 369 p. ; 21 cm. – (Vocabularios temáticos ; 4). - D.L. C 2458-2008. – ISBN 978-84-9887-018-3 1.Medicina �������������������������������������������������������������������������veterinaria-Diccionarios�������������������������������������������������. 2.Galego (Lingua)-Glosarios, vocabularios, etc. políglotas. I.Lombardero Fernández, Matilde. II.Rodríguez Rio, Xusto A. coord. III. Universidade de Santiago de Compostela. Servizo de Normalización Lingüística, coord. IV.Universidade de Santiago de Compostela. Servizo de Publicacións e Intercambio Científico, ed. V.Serie. 591.4(038)=699=60=20 Coordinador Xusto A. Rodríguez Río (Área de Terminoloxía. Servizo de Normalización Lingüística. Universidade de Santiago de Compostela) Autoras/res Matilde Lombardero Fernández (doutora en Veterinaria e profesora do Departamento de Anatomía e Produción Animal. -
Avian Adrenal Medulla: Cytomorphology and Function
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Publications of the IAS Fellows Volume 45(1-4):1-11, 2001 Acta Biologica Szegediensis http://www.sci.u-szeged.hu/ABS REVIEW ARTICLE Avian adrenal medulla: cytomorphology and function Asok Ghosh, Stephen W. Carmichael1*, Monisha Mukherjee Department of Zoology, University of Calcutta, Calcutta, India, 1Department of Anatomy, Mayo Clinic/Foundation, Rochester, Minnesota, USA ABSTRACT The purpose of this review is to explore the world literature on the avian adrenal KEY WORDS medulla from the last 20 years. Unlike the mammalian adrenal medulla, the adrenal gland in adrenal medulla birds has chromaffin cells mixed with cortical cells. Studies have investigated the ultrastructure birds (both transmission and scanning electron microscopy), biochemistry, and physiology (partic- morphology ularly interactions with other endocrine glands) of the avian adrenal medulla. Although function progress has been made, it is apparent that research on the avian adrenal medulla still lags behind work on the mammalian organ. Acta Biol Szeged 45(1-4):1-11 (2001) The adrenal glands of birds, like those in mammals, are in the adrenal medulla. This profound variation of medullary paired yellow- or orange-colored pear- or triangle-shaped E/NE ratio in birds suggests a distinct evolutionary pattern glands that are next to the kidneys. The intermingling nature (Ghosh 1977, 1980). The avian phylogeny used in this study of cortical and medullary components constitutes a major was essentially based on palaeontological evidences (Grego- characteristic of avian adrenal medulla (Vestergaard and ry 1957). We feel that our “claim” of hormonal taxonomy is Willeberg 1978). -

Endocrine Tumors – Adrenal Medulla
Endocrine Tumors – Adrenal Medulla 803-808-7387 www.gracepets.com These notes are provided to help you understand the diagnosis or possible diagnosis of cancer in your pet. For general information on cancer in pets ask for our handout “What is Cancer”. Your veterinarian may suggest certain tests to help confirm or eliminate diagnosis, and to help assess treatment options and likely outcomes. Because individual situations and responses vary, and because cancers often behave unpredictably, science can only give us a guide. However, information and understanding for tumors in animals is improving all the time. We understand that this can be a very worrying time. We apologize for the need to use some technical language. If you have any questions please do not hesitate to ask us. What are the adrenal glands? The adrenal glands are located close to the kidneys. They are part of the body’s endocrine system, the glands of which also include the pituitary (in the brain), thyroid, parathyroid and Islets of Langerhans in the pancreas. Endocrine glands produce specialized chemicals called “hormones”. These regulate and integrate many activities to maintain internal stability of the body. The hormones pass directly into the blood to affect target cells elsewhere. Hormones are also produced by many cells in other tissues. Each of the two adrenal glands has two parts. The outer part (cortex) is controlled by a hormone (adrenocorticotrophic hormone, ACTH) from the pituitary gland. The cortex produces steroid hormones of several types. The inner part (medulla) of the adrenal gland originates from the same cells that in the embryo form the nervous system, and therefore not surprisingly it produces neuroendocrine hormones with effects similar to those of the sympathetic nervous system. -

Chapter 20: Endocrine System
EndocrineEndocrine SystemSystem Modified by M. Myers 1 TheThe EndocrineEndocrine SystemSystem 2 EndocrineEndocrine GlandsGlands z The endocrine system is made of glands & tissues that secrete hormones. z Hormones are chemicals messengers influencing a. metabolism of cells b. growth and development c. reproduction, d. homeostasis. 3 HormonesHormones Hormones (chemical messengers) secreted into the bloodstream and transported by blood to specific cells (target cells) Hormones are classified as 1. proteins (peptides) 2. Steroids 4 HormoneHormone ClassificationClassification z Steroid Hormones: – Lipid soluble – Diffuse through cell membranes – Endocrine organs z Adrenal cortex z Ovaries z Testes z placenta 5 HormoneHormone ClassificationClassification z Nonsteroid Hormones: – Not lipid soluble – Received by receptors external to the cell membrane – Endocrine organs z Thyroid gland z Parathyroid gland z Adrenal medulla z Pituitary gland z pancreas 6 HormoneHormone ActionsActions z “Lock and Key” approach: describes the interaction between the hormone and its specific receptor. – Receptors for nonsteroid hormones are located on the cell membrane – Receptors for steroid hormones are found in the cell’s cytoplasm or in its nucleus 7 http://www.wisc- online.com/objects/index_tj.asp?objID=AP13704 8 EndocrineEndocrine SystemSystem z There is a close assoc. b/w the endocrine & nervous systems. z Hormone secretion is usually controlled by either negative feedback or antagonistic hormones that oppose each other’s actions 9 HypothalamusHypothalamus 1. regulates the internal environment through the autonomic system 2. controls the secretions of the pituitary gland. 10 HypothalamusHypothalamus && PituitaryPituitary GlandGland posteriorposterior pituitary/pituitary/ anterioranterior pituitarypituitary 11 PosteriorPosterior PituitaryPituitary The posterior pituitary secretes zantidiuretic hormone (ADH) zoxytocin 12 13 14 AnteriorAnterior pituitarypituitary glandgland 1. -

The Endocrine System
4/12/2016 Adrenal Glands Image From: http://www.hawaiilife.com/articles/2012/03/good-news-vacation-rental-owners/ 70 Copyright © 2009 Pearson Education, Inc Figure 10.14a Adrenal Glands Adrenal Adrenal cortex Adrenal Glands gland • Mineralocorticoids • Gonadocorticoids • Glucocorticoids • Controlled by both nerves and hormones – Adrenal medulla Adrenal Medulla • Epinephrine • Controlled by nerves from the hypothalamus • Norepinephrine – Adrenal Cortex • Controlled by ACTH (a hormone) secreted by the anterior pituitary gland (b) A section through the adrenal gland reveals two regions, the outer adrenal cortex and the inner adrenal medulla. These regions secrete different hormones. 73 Copyright © 2009 Pearson Education, Inc Figure 10.14b Adrenal Glands Adrenal Glands • Adrenal Medulla • Adrenal Cortex – Epinephrine – 2 types of hormones secreted • Increases blood pressure • Mineralocorticoids • Increases heart rate • Glucocorticoids • Increases blood glucose levels 74 Copyright © 2009 Pearson Education, Inc 75 Copyright © 2009 Pearson Education, Inc 1 4/12/2016 Adrenal Glands - Cortex Adrenal Glands - Cortex • Mineralocorticoids • Aldosterone – Example: Aldosterone – Promotes renal absorption of Na+ and renal – Mineral homeostasis excretion of K+ – Water balance – Increased blood pressure • Target – Kidneys 76 Copyright © 2009 Pearson Education, Inc 77 Copyright © 2009 Pearson Education, Inc Adrenal Glands - Cortex Adrenal Glands - Cortex • Glucocorticoids • Cortisol – Example: Cortisol – Effects glucose homeostasis – Act on the liver to -

Adrenal and Paraganglia Tumors Adrenal Cortical Tumors IHC: (+) SF1, Inhibin, Melan-A, Calretinin, Synaptophysin, (-) Chromogranin, Cytokeratin, S100
Last updated: 11/11/2020 Prepared by Kurt Schaberg Adrenal and Paraganglia Tumors Adrenal Cortical Tumors IHC: (+) SF1, Inhibin, Melan-A, Calretinin, Synaptophysin, (-) Chromogranin, Cytokeratin, S100. Often variable!! Adrenal Cortical Adenoma Benign. Very common. Often incidentally identified. Usually unilateral solitary masses with atrophic background adrenal gland. Tumor cells can be lipid-rich (clearer) or lipid-poor (pinker) arranged in nests and cords separated by abundant vasculature. Occasional lipofuscin pigment. Nuclei generally small and round (occasional extreme “endocrine atypia” is common). Low/no mitotic activity. Intact reticulin framework. On a spectrum with and may hard to differentiate from hyperplastic nodules, which is more often multinodular (background hyperplasia) and bilateral. Can be non-functional (85%) or functional (15%). Associated with MEN1, FAP, Carney Complex, among Aldosterone-producing→ “Conn syndrome”→ others… hypertension and hypokalemia If aldosterone-secreting adenoma is treated with Cortisol-producing→ (ACTH-independent) spironolactone→ “spironolactone bodies” (below) “Cushing Syndrome” → central obesity, moon face, hirsutism, poor healing, striae Sex-hormone-producing→ Rare (more common in carcinomas). Symptoms depend on hormone/sex (virilization or feminization) Adrenal Cortical Carcinoma Malignant. Most common in older adults. Can present with an incidental unilateral mass or with an endocrinopathy (see above). Solid, broad trabeculae, or large nested growth (more diffuse, and larger groups than in adenomas) Thick fibrous capsule with occasional fibrous bands. Frequent tumor necrosis. Frequent vascular or capsular invasion. Increased mitotic activity. Variants: Oncocytic, Myxoid, Sarcomatoid Mostly sporadic, but can be associated with Lynch Syndrome and Li-Fraumeni Syndrome Distinguishing between an Adrenal Cortical Adenoma vs Carcinoma Weiss Criteria: Weiss Criteria (≥3 = Malignant) Most widely used system, but doesn’t work as well High nuclear grade (based of Fuhrman criteria) in borderline cases or variants. -

Adrenal Medulla
Adrenal medulla Objectives: ❖ Summarize the actions of adrenal androgens. ❖ Describe the causes and major manifestations of hyperadrenocorticism and Hypoadrenocorticism. ❖ Describe circumstances in which catecholamines are released from the adrenal gland. ❖ List the major actions of catecholamines. Done by : ➔ Team leader: Rahaf AlShammari, Abdulelah AlDossari ➔ Team members: ◆ Esraa AlNazzawi, Renad AlMogren ◆ Abdulmajeed AlWardi, Abdulelah AlSaeed Colour index: ◆ Anas AlSaif, Saif AlMeshari ● Important ◆ Laila AlSabbagh, Fatimah Blasharaf ● ;) ◆ Renad AlSwailmy, Wejdan AlShamery ● Extra َوأَن ﻟﱠﯾْسَ ﻟِ ِْﻺَﻧﺳِﺎن إِﱠﻻ ﻣَﺎ َﺳَﻌٰﻰ Adrenal medulla ● The adrenal medulla is the inner part or core of each adrenal gland. ● It is considered as part of sympathetic nervous system. ● It synthesizes and secretes catecholamines from Tyrosine: Adrenaline (epinephrine) -- 80% of the secretion. Noradrenaline (norepinephrine) -- 20 % of the secretion. Small amount of dopamine ● They are released from chromaffin cells ● Secretion of these hormones causes: Blood to be diverted to the brain, heart, and skeletal muscle. ● Epinephrine is the more potent stimulator of the heart and metabolic activities ● Norepinephrine is more influential on peripheral vasoconstriction and blood pressure ● High levels of cortisol that drain into the medulla from the adrenal cortex induce expression of the enzyme phenylethanolamine N-methyltransferase (PNMT), which converts norepinephrine to epinephrine Role of the adrenal medullary hormones 1. Enhance the effects of the 2. Prepare -
The Endocrine System
PowerPoint® Lecture Slides The Endocrine System: An Overview prepared by Leslie Hendon University of Alabama, Birmingham • A system of ductless glands • Secrete messenger molecules called hormones C H A P T E R 17 • Interacts closely with the nervous system Part 1 • Endocrinology The Endocrine • Study of hormones and endocrine glands System Copyright © 2011 Pearson Education, Inc. Copyright © 2011 Pearson Education, Inc. Endocrine Organs Location of the Major Endocrine Glands Pineal gland • Scattered throughout the body Hypothalamus Pituitary gland • Pure endocrine organs are the … Thyroid gland • Pituitary, pineal, thyroid, parathyroid, and adrenal Parathyroid glands glands (on dorsal aspect of thyroid gland) • Organs containing endocrine cells include: Thymus • Pancreas, thymus, gonads, and the hypothalamus Adrenal glands • Plus other organs secrete hormones (eg., kidney, stomach, intestine) Pancreas • Hypothalamus is a neuroendocrine organ • Produces hormones and has nervous functions Ovary (female) Endocrine cells are of epithelial origin • Testis (male) Copyright © 2011 Pearson Education, Inc. Copyright © 2011 Pearson Education, Inc. Figure 17.1 Hormones Control of Hormones Secretion • Classes of hormones • Amino acid–based hormones • Secretion triggered by three major types of • Steroids—derived from cholesterol stimuli: • Basic hormone action • Humoral—simplest of endocrine control mechanisms • Circulate throughout the body in blood vessels • Secretion in direct response to changing • Influences only specific tissues— those with ion or nutrient levels in the blood target cells that have receptor molecules for that hormone • Example: Parathyroid monitors calcium • A hormone can have different effects on • Responds to decline by secreting different target cells (depends on the hormone to reverse decline receptor) Copyright © 2011 Pearson Education, Inc. Copyright © 2011 Pearson Education, Inc. -

Comparison of the Anterior Pituitary-Adrenal Cortical Stimulating Effect of U.S.P. Epinephrine, Synthetic L-Epinephrine, and Nor-Epinephrine
COMPARISON OF THE ANTERIOR PITUITARY-ADRENAL CORTICAL STIMULATING EFFECT OF U.S.P. EPINEPHRINE, SYNTHETIC L-EPINEPHRINE, AND NOR-EPINEPHRINE Leonard L. Madison J Clin Invest. 1950;29(6):789-791. https://doi.org/10.1172/JCI102316. Research Article Find the latest version: https://jci.me/102316/pdf COMPARISON OF THE ANTERIOR PITUITARY-ADRENAL CORTICAL STIMULATING EFFECT OF U.S.P. EPI- NEPHRINE, SYNTHETIC L-EPINEPHRINE, AND NOR-EPINEPHRINE 1 BY LEONARD L. MADISON (From the Department of Medicine, Southwestern Medical School of the University of Texas, and the Department of Medicine of the Veterans Administration Hospital, Dallas, Texas) (Submitted for publication December 16, 1949; accepted, March 6, 1950) The recent demonstration of the dramatic thera- adrenal cortical steroid hormone production with peutic effectiveness of the adrenal 11, 17-oxy- resultant eosinopenia and increased urinary 11- steroids (Compound E of Kendall) in the therapy oxysteroid and 17-ketosteroid excretion. of rheumatoid arthritis, rheumatic fever, and lu- Although long suspected, it has only recently pus erythematosus has lent great impetus to the been proven that the adrenal medulla secretes two study of the role of the adrenal cortex in these hormones, epinephrine and its non-methylated mesenchymal diseases, and to the search for new analogue, nor-epinephrine (arterenol, 1- [3, 4- methods of stimulating the adrenal to produce dehydroxyphenyl] 2-aminoethanol) (7, 8). As- endogenously these therapeutic agents in greater says on U. S. P. epinephrine reference standards, abundance (1-3). The wide variety of stimuli and U. S. P. commercial epinephrine revealed a capable of producing an "alarm reaction" are all nor-epinephrine content varying from 10 to 35%, attended by one common factor, the stimulation of the average being 15%o (7-9). -

Adrenal & Gonadal Hormones Layers of Adrenal Cortex
Adrenal & Gonadal Hormones Topics for today: •Adrenal cortex hormone •Adrenal medulla hormones •Hormone control of organs •Steroid hormone synthesis •Vitamin D3 • Estrogens and Progesterone Layers of adrenal cortex zona glomerulosa zona fasiculata zona reticularis 1 Hormones of adrenal cortex zona glomerulosa zona fasiculata O zona reticularis HO dehydroepiandrosterone Androgens – dehydroepiandrosterone • increased protein synthesis DHEA is weak androgen with • masculinizing effects in almost no effect in male but has female (hypersecretion) masculinizing effects in females Adrenal medulla (interior) • Composed of modified post-synaptic sympathetic neurons • Releases mostly epinephrine. • Has effects similar to those triggered by sympathetic nervous system Adrenal medulla hormones HO HO -CH-CH2 - N-CH3 epinephrine OH H Effects of epinephrine: • causes elevated blood glucose level • stimulates glycolysis & fatty acid use • increases cardiac output & blood pres • shifts blood flow to skeletal muscle • increases rate and depth of respiration 2 Organ responses to Epinephrine Causes glycogen Causes fatty acid release degradation in muscle from adipose tissue fatty glycogen acids trigly- cerides lactate glycerol muscle lactate adipose tissue Causes release of glycogeno lysis glucose from liver glucose ...elevated plasma level liver Hormone action time Epinephrine is in group of fast-acting hormones Fast-acting Hormones Slow-acting Hormones • Norepinephrine • Throxine • Epinephrine • Cortisol • Insulin • Growth hormone • glucagon • Estrogens -
![Endocrine Glands [PDF]](https://docslib.b-cdn.net/cover/5879/endocrine-glands-pdf-3555879.webp)
Endocrine Glands [PDF]
Histology of Skin and Endocrine glands Skin and Endocrine glands • Skin • Thyroid • Parathyroid gland • Adrenal gland • Pituitary gland • Pineal gland Skin • Layers of skin • Epidermis • Five layers • Dermis • Two layers Junction • Dermal papilla • Epidermal peg (rete pegs) Skin…. • Epidermis - 1.Stratum basale • Single layer of columnar cells 2.Stratum spinosum • Several layers of polyhedral cells, spine like process, tonofilament 3.Stratum granulosum • Keratohyline granules 4.Stratum lucidum • Homogeneous keratin, fusiform cells 5.Stratum corneum-non nucleated keratinized dead cells Skin…… • Cells of epidermis -Keratinocytes- 90%,able to keratinization -Cells of Langherhans- present in st.spinosum, clear cytoplasmic process, antigen producing cell -Melamocytes-pigmented cell in basal layer, many cytoplasmic process. Produce Melanin -Merkel cells- sensory cell Skin….. Dermis • Papillary layer • Tactile papilla • Vascular papilla • Collagen fibre • Reticular layer Collagen fibre • Sweat glands • Sebaceous glands • Hairs Skin…… • Thick skin • Thin skin Thyroid gland 1.Capsule 2.Parenchyma • thyroid follicle -Structural & functional unite -Epithelium-simple cuboidal cells (follicular cells), synthesis thyroxin hormone -cell size varies with activeness -Lumen of thyroid follicle filled with colloid -Parafollicular cells (“C” Cells) Present at margin or inter follicular space, Calcitonin 3.Stroma- connective tissue, septa, blood vessels Parathyroid gland • Chief cells • Polygonal shape, round nucleus, synthesis Parathormone • Oxyphill cells-