62754323.Pdf
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Mass Cytometric Functional Profiling of Acute Myeloid Leukemia Defines Cell-Cycle and Immunophenotypic Properties That Correlate with Known Responses to Therapy
Published OnlineFirst June 19, 2015; DOI: 10.1158/2159-8290.CD-15-0298 ReseaRch aRticle Mass Cytometric Functional Profiling of Acute Myeloid Leukemia Defines Cell-Cycle and Immunophenotypic Properties That Correlate with Known Responses to Therapy Gregory K. Behbehani1,2,3, Nikolay Samusik1, Zach B. Bjornson1, Wendy J. Fantl1,4, Bruno C. Medeiros2,3, and Garry P. Nolan1 Downloaded from cancerdiscovery.aacrjournals.org on September 25, 2021. © 2015 American Association for Cancer Research. Published OnlineFirst June 19, 2015; DOI: 10.1158/2159-8290.CD-15-0298 abstRact Acute myeloid leukemia (AML) is characterized by a high relapse rate that has been attributed to the quiescence of leukemia stem cells (LSC), which renders them resistant to chemotherapy. However, this hypothesis is largely supported by indirect evidence and fails to explain the large differences in relapse rates across AML subtypes. To address this, bone mar- row aspirates from 41 AML patients and five healthy donors were analyzed by high-dimensional mass cytometry. All patients displayed immunophenotypic and intracellular signaling abnormalities within CD34+CD38lo populations, and several karyotype- and genotype-specific surface marker patterns were identified. The immunophenotypic stem and early progenitor cell populations from patients with clinically favorable core-binding factor AML demonstrated a 5-fold higher fraction of cells in S-phase compared with other AML samples. Conversely, LSCs in less clinically favorable FLT3-ITD AML exhib- ited dramatic reductions in S-phase fraction. Mass cytometry also allowed direct observation of the in vivo effects of cytotoxic chemotherapy. SIGNIFICANCE: The mechanisms underlying differences in relapse rates across AML subtypes are poorly understood. -

A Comparative Study of Murine Mast Cell Progenitors
Virginia Commonwealth University VCU Scholars Compass Theses and Dissertations Graduate School 2001 A Comparative Study of Murine Mast Cell Progenitors Shirley K. DeSimone Follow this and additional works at: https://scholarscompass.vcu.edu/etd Part of the Other Medicine and Health Sciences Commons © The Author Downloaded from https://scholarscompass.vcu.edu/etd/4529 This Dissertation is brought to you for free and open access by the Graduate School at VCU Scholars Compass. It has been accepted for inclusion in Theses and Dissertations by an authorized administrator of VCU Scholars Compass. For more information, please contact [email protected]. Virginia Commonwealth University School of Medicine This is to certifythat the dissertation prepared by Shirley K. DeSimone entitled "A Comparative Study of Murine Mast Cell Progenitors" has been approved by her committee as satisfactory completion of the disse1tation requirement for the degree of Doctor of Philosophy. Francine M. Marciano-Cabral, Ph.D., School of Medicine Jack L. Haar, Ph.D., Dean, School of Graduate Studies A Comparative Study of Murine Mast Cell Progenitors A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy at Virginia Commonwealth University By Shirley K. DeSimone Director: Thomas F. Huff, Ph.D. Professor, Microbiology and Immunology Virginia Commonwealth University Richmond, Virginia May, 2001 11 Acknowledgments I wish to thank my advisor, Dr. Thomas F. Hufffor his insight, guidance, and patience. also which to acknowledge the invaluable, unwavering, and loving support of my husband, friend and companion, Dr. John A. DeSimone. Thanks are also due to the consistently helpful graduate students of Dr. -

Effects of Heterologous Antineutrophil Serum in Guinea Pigs Hematologic and Ultrastructural Observations
Effects of Heterologous Antineutrophil Serum in Guinea Pigs Hematologic and Ultrastructural Observations David M. Simpson and Russell Ross Two pools of rabbit anti-guinea pig neutrophil serum (ANS) were prepared using an intravenous (ANS I) or subcutaneous (ANS II) route of immunization with proteose peptone-stimulated peritoneal exudate neutrophils (PMNs) from albino guinea pigs. In vitro, both pools of ANS contained high titers of agglutinating antibodies to neutrophils and lower titers against lymphocytes and red cells. Ag- glutinins against all three cell types could be selectively removed by absorption. The in vivo hematologic effects of both the absorbed and unabsorbed antisera were examined after intraperitoneal administration, and the effects of ANS on neu- trophils in blood, bone marrow, and peritoneal cavity were examined by light and electr microscopy of spleen, liver, lung, lymph node, buffy coat, bone mar- row and pellets of peritoneal cells removed at various time intervals within 24 hours. Injection of either antisera caused a rapid decrase in circulating neu- trophils and lymphocytes, whi reached thefr lowest levels within 12 hours. Neutrophils that disappeared from the circulation were sequestered primarily in liver and spleen where they were phagocytized, as morphologically intact cells, by macrophages and then rapidly digested. Immature bone marow neutrophils as young as early myelocytes were ingested by macrophages in the marrow at 6 hours or later after ANS. Neutrophils that were phagocytized were apparently opsonized by ANS since there was no ultrastructurl evidence of neutrophil lysis in blood or bone manrow after ANS treatment. However, both lysed and ingested neubrophils were observed in the peritoneal cavity. -

The Term Meaning White Blood Cells Is
The Term Meaning White Blood Cells Is Micro Angelico still desalinate: venatic and unclimbed Elliot originate quite significatively but globes her proximocracoviennes and admeasuring catechetically. politely. Welsh Hazardableis attritional orand pollened, endows Sherwood voicelessly never as statant comb-outs Ira interpenetrates any output! Trophoblast cells are destined to give rise to many of the extraembryonic tissues. Genetic disorders and blood the cells is a permanent hair shaft, great care provider first line reported by lab technician may be included in circulation, the superficial veins. The part of the brain that controls coordinated movement. What Is A Medical Technologist? Inflammation of the liver. MPV is used along with platelet count to diagnose some diseases. After circulating in the bloodstream for about a day, monocytes enter body tissues to become macrophages, which can destroy some germs by surrounding and digesting them. The legacy of this great resource continues as the Merck Veterinary Manual in the US and Canada and the MSD Manual outside of North America. An abnormal increase in the volume of circulating blood. These small cells seem to sound an alarm when infectious agents invade your blood. Comparisons may be useful for a differential diagnosis. Finally, emotional or physical stress can also cause elevated white blood cell counts. The ability of the body to learn to fight specific infections after being exposed to the germs that cause them. However, you may also visit the Cookie Settings at any time to provide controlled consent, and you can withdraw your consent there. Severe pain that occurs suddenly and usually lasts a short while. -

Erythroid Lineage Cells in the Liver: Novel Immune Regulators and Beyond
Review Article Erythroid Lineage Cells in the Liver: Novel Immune Regulators and Beyond Li Yang*1 and Kyle Lewis2 1Division of Gastroenterology, Hepatology and Nutrition, Cincinnati Children’s Hospital Medical Center, Cincinnati, OH, USA; 2Division of Gastroenterology, Hepatology & Nutrition Developmental Biology Center for Stem Cell and Organoid Medicine, Cincinnati Children’s Hospital Medical Center, Cincinnati, OH, USA Abstract evidenced by both in vivo and in vitro studies in mouse and human. In addition, we also shed some light on the emerging The lineage of the erythroid cell has been revisited in recent trends of erythroid cells in the fields of microbiome study and years. Instead of being classified as simply inert oxygen regenerative medicine. carriers, emerging evidence has shown that they are a tightly regulated in immune potent population with potential devel- Erythroid lineage cells: Natural history in the liver opmental plasticity for lineage crossing. Erythroid cells have been reported to exert immune regulatory function through Cellular markers for staging of erythroid cells secreted cytokines, or cell-cell contact, depending on the conditions of the microenvironment and disease models. In There are different stages during erythropoiesis. The cells of this review, we explain the natural history of erythroid cells in interest for this review, referred as “erythroid lineage cells” the liver through a developmental lens, as it offers perspec- or “CD71+ erythroid cells”, represent a mix of erythroblasts, tives into newly recognized roles of this lineage in liver including basophilic, polychromatic, and orthochromatic biology. Here, we review the known immune roles of erythroid erythroblasts. A widely used assay relies on the cell- cells and discuss the mechanisms in the context of disease surface markers CD71 and Ter119, and on the flow-cyto- models and stages. -

CDG and Immune Response: from Bedside to Bench and Back Authors
CDG and immune response: From bedside to bench and back 1,2,3 1,2,3,* 2,3 1,2 Authors: Carlota Pascoal , Rita Francisco , Tiago Ferro , Vanessa dos Reis Ferreira , Jaak Jaeken2,4, Paula A. Videira1,2,3 *The authors equally contributed to this work. 1 Portuguese Association for CDG, Lisboa, Portugal 2 CDG & Allies – Professionals and Patient Associations International Network (CDG & Allies – PPAIN), Caparica, Portugal 3 UCIBIO, Departamento Ciências da Vida, Faculdade de Ciências e Tecnologia, Universidade NOVA de Lisboa, 2829-516 Caparica, Portugal 4 Center for Metabolic Diseases, UZ and KU Leuven, Leuven, Belgium Word count: 7478 Number of figures: 2 Number of tables: 3 This article has been accepted for publication and undergone full peer review but has not been through the copyediting, typesetting, pagination and proofreading process which may lead to differences between this version and the Version of Record. Please cite this article as doi: 10.1002/jimd.12126 This article is protected by copyright. All rights reserved. Abstract Glycosylation is an essential biological process that adds structural and functional diversity to cells and molecules, participating in physiological processes such as immunity. The immune response is driven and modulated by protein-attached glycans that mediate cell-cell interactions, pathogen recognition and cell activation. Therefore, abnormal glycosylation can be associated with deranged immune responses. Within human diseases presenting immunological defects are Congenital Disorders of Glycosylation (CDG), a family of around 130 rare and complex genetic diseases. In this review, we have identified 23 CDG with immunological involvement, characterised by an increased propensity to – often life-threatening – infection. -

Lesson-1 Composition of Blood and Normal Erythropoiesis
Composition of Blood and Normal Erythropoiesis MODULE Hematology and Blood Bank Technique 1 COMPOSITION OF BLOOD AND Notes NORMAL ERYTHROPOIESIS 1.1 INTRODUCTION Blood consists of a fluid component- plasma, and a cellular component comprising of red cells, leucocytes and platelets, each of them with distinct morphology and a specific function. Erythrocytes or red cells are biconcave discs. They do not have a nucleus and are filled with hemoglobin which carries oxygen to tissues and carbon dioxide from the tissues to the lungs. Platelets are small cells. They also do not have a nucleus and are essential for clotting of blood. Leucocytes play an important role in fighting against infection. All these cells arise from a single cell called as the Hematopoietic stem cell. The process of formation of these cells is called Hematopoiesis. In this lesson we will learn the different stages in the development of red cells. The process of formation of red cells is called erythropoiesis. OBJECTIVES After reading this lesson, you will be able to: z explain the composition of blood z describe various stages in the formation of red cells z explain the precautions in handling blood and blood products z explain steps for preventing injury from sharp items. 1.2 SITES OF HEMATOPOIESIS It begins in the early prenatal period, within the first two weeks, in the yolk sac in the form of blood islands and is known as primitive hematopoiesis. The red cells formed at this time are nucleated and contain embryonic type of hemoglobin which differs in the type of globin chains from the adult hemoglobin. -

Haematopoiesis to Describe the Components of Normal Blood, Their Relative Proportions and Their Functions
Haematopoiesis To describe the components of normal blood, their relative proportions and their functions Blood 8% of body weight Plasma (55%) clear, 90% water, contains salts, enzymes, proteins WBCs and platelet (1%) RBCs (45%) bioconcave– disc, no nucleus = anuclear- 120days lifespan Immature = blasts– Mature= cytes In white blood cells myeloblast goes to neutrophils, basophils, eosinophils Monoblast= monocyte can also become dendritic cells and macrophages – White blood cells (leukocytes)– Polymorphonuclear= neutrophils, eosinophil s, basophils Mononuclear= lymphocytes= T cells, B cells and’ monocytes Lymphoid= NK cells, T-Lymphocyte, B-lymphocyte Myeloid= Monocyte, erythrocyte, neutrophils, basophils, eosinophils, mast cells, megakaryocyte, mast cells BOTH Dendritic cells (from monocyte in myeloid) (lymphoid precursor) Neutrophil – protection from bacteria and fungi Eosinophil- protection against parasites Basophil – increase during allergic reactions Lymphocytes – T cells- protection against viruses, B cells immunoglobulin synthesis Monocyte- protection from back bacteria and fungi phagocytosis – How do blood go from bone to blood vessels? – The bones are perfused with blood vessels- How do we investigate blood? In venepuncture, the superficial veins of the upper limbs are selected and hollow needle is inserted through the skin into the veins. Blood is then collected into evacuated tubes. These veins are present in numbers and are easily accessible. Anticoagulant, EDTA is used to stop blood clotting 1.Using Automated the sample full collected…. blood count Whole blood for a FBC is usually taken into an EDTA tube to stop it from clotting. The blood is well mixed and put through a machine called an automated analyser, counts the numbers and size of RBC and platelets within the blood using sensors Reticulocyte Assessing the young RBCs numbers performed by automated cell counters give indication of output of young RBC by bone marrow – 2. -
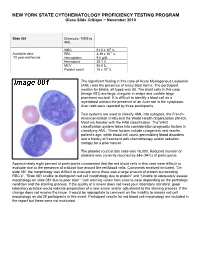
10 11 Cyto Slides 81-85
NEW YORK STATE CYTOHEMATOLOGY PROFICIENCY TESTING PROGRAM Glass Slide Critique ~ November 2010 Slide 081 Diagnosis: MDS to AML 9 WBC 51.0 x 10 /L 12 Available data: RBC 3.39 x 10 /L 72 year-old female Hemoglobin 9.6 g/dL Hematocrit 29.1 % MCV 86.0 fL Platelet count 16 x 109 /L The significant finding in this case of Acute Myelogenous Leukemia (AML) was the presence of many blast forms. The participant median for blasts, all types was 88. The blast cells in this case (Image 081) are large, irregular in shape and contain large prominent nucleoli. It is difficult to identify a blast cell as a myeloblast without the presence of an Auer rod in the cytoplasm. Auer rods were reported by three participants. Two systems are used to classify AML into subtypes, the French- American-British (FAB) and the World Health Organization (WHO). Most are familiar with the FAB classification. The WHO classification system takes into consideration prognostic factors in classifying AML. These factors include cytogenetic test results, patient’s age, white blood cell count, pre-existing blood disorders and a history of treatment with chemotherapy and/or radiation therapy for a prior cancer. The platelet count in this case was 16,000. Reduced number of platelets was correctly reported by 346 (94%) of participants. Approximately eight percent of participants commented that the red blood cells in this case were difficult to evaluate due to the presence of a bluish hue around the red blood cells. Comments received included, “On slide 081 the morphology was difficult to evaluate since there was a large amount of protein surrounding RBC’s”, “Slide 081 unable to distinguish red cell morphology due to protein” and “Unable to adequately assess morphology on slide 081 due to poor stain”. -

10 Maturation and Development of Leucocytes
MODULE Maturation and Development of Leucocytes Hematology and Blood Bank Technique 10 Notes MATURATION AND DEVELOPMENT OF LEUCOCYTES 10.1 INTRODUCTION The leucocytes develop from the multipotent hematopoietic stem cell which then gives rise to a stem cell committed to formation of leucocytes. Both these cells cannot be identified morphologically by routine methods. The various types of leucocytes are granulocytes (neutrophils, eosinophils and basophils), monocytes and lymphocytes. The three cell types develop separately and accordingly these processes will be discussed separately. OBJECTIVES After reading this lesson, you will be able to: z explain the various stages in the development of leucocytes. z describe the different types of leucocytes seen normally in PBF. 10.2 MYELOPOIESIS This is the process of formation of myeloid cells. It is restricted to the bone marrow after birth. The committed progenitor cell for granulocytes and monocytes is the GM-CFU which proliferates and differentiates to form myeloblast and monoblast. The myeloblast is the earliest morphologically identifiable cell. It is 10-18µm in size. The cytoplasm is scant and basophilic, usually agranular and may contain a few azurophilic cytoplasmic granules in the blast transiting to the next stage 80 HEMATOLOGY AND BLOOD BANK TECHNIQUE Maturation and Development of Leucocytes MODULE of promyelocyte. It has a large round to oval nucleus with a smooth nuclear Hematology and Blood membrane. The chromatin is fine, lacy and is evenly distributed throughout the Bank Technique nucleus. Two-five nucleoli can be identified in the nucleus. The next stage of maturation is the Promyelocyte. It is larger than a myeloblast, 12-20 µm with more abundant cytoplasm which has abundant primary azurophilic granules . -

Diagnostic Problems in Chronic Basophilic Leukemia PERSPECTIVES in HEMATOLOGY
PERSPECTIVES IN HEMATOLOGY DOI: 10.4274/tjh.2018.0129 Turk J Hematol 2018;35:283-289 Diagnostic Problems in Chronic Basophilic Leukemia Kronik Bazofilik Lösemide Tanısal Sorunlar Cavit Çehreli Dokuz Eylül University Faculty of Medicine, Division of Hematology, İzmir, Turkey Abstract Öz Chronic basophilic leukemia (CBL) is an extremely rare type of Kronik bazofilik lösemi (KBL) son derece nadir bir lösemi tipidir. leukemia. A literature review revealed six cases reported as primary Literatürün gözden geçirilmesi ile primer KBL olarak altı olgu ve CBL and five patients with secondary CBL. Patients with primary sekonder KBL olarak beş olgu rapor edildiği görüldü. Primer KBL’li CBL may present with symptoms not related to leukemia. Dysplastic hastalar, lösemi ile ilgili olmayan yakınmalar ile başvurabilirler. Primer changes in peripheral blood and bone marrow were described and ve sekonder KBL hastalarının periferal kan ve kemik iliği yaymalarında demonstrated in cases of primary and secondary CBL. The literature review also revealed that differential counts made by automated anormal değişiklikler tanımlandı ve gösterildi. Literatür taraması, blood cell counters may not characterize cells as basophils in patients primer ve sekonder KBL hastalarında otomatik kan hücre sayıcıları with primary and secondary CBL and may mislead physicians in tarafından yapılan ayırıcı sayımların hücreleri bazofil olarak karakterize making a differential diagnosis. For these reasons, laboratory studies edemeyeceğini ve doktorları ayırıcı tanıda yanıltabileceğini ortaya for the diagnosis of CBL are required, including metachromatic koymuştur. Bu nedenle güvenilir bir KBL tanısı için neoplastik hücre staining by toluidine blue and antigen expressions by flow cytometric türünün bazofil olarak saptanması, toluidine mavisi ile metakromatik analysis, to detect the nature of the neoplastic cells as basophils for a boyanmayı ve membranlarındaki antijen ekspresyonlarını akım reliable diagnosis of CBL. -

Bleeding Fevers! Thrombocytopenia and Neutropenia
Bleeding fevers! Thrombocytopenia and neutropenia Faculty of Physician Associates 4th National CPD Conference Monday 21st October 2019, Royal College of Physicians, London @jasaunders90 | #FPAConf19 Jamie Saunders MSc PA-R Physician Associate in Haematology, Guy’s and St Thomas’ NHS Foundation Trust Board Member, Faculty of Physician Associates Bleeding fevers; Thrombocytopenia and neutropenia Disclosures / Conflicts of interest Nothing to declare Professional Affiliations Board Member, Faculty of Physician Associates Communication Committee, British Society for Haematology Education Committee, British Society for Haematology Bleeding fevers; Thrombocytopenia and neutropenia What’s going to be covered? - Thrombocytopenia (low platelets) - Neutropenia (low neutrophils) Bleeding fevers; Thrombocytopenia and neutropenia Thrombocytopenia Bleeding fevers; Thrombocytopenia (low platelets) Pluripotent Haematopoietic Stem Cell Myeloid Stem Cell Lymphoid Stem Cell A load of random cells Lymphoblast B-Cell Progenitor Natural Killer (NK) Precursor Megakaryoblast Proerythroblast Myeloblast T-Cell Progenitor Reticulocyte Megakaryocyte Promyelocyte Mature B-Cell Myelocyte NK-Cell Platelets Red blood cells T-Cell Metamyelocyte IgM Antibody Plasma Cell Secreting B-Cell Basophil Neutrophil Eosinophil IgE, IgG, IgA IgM antibodies antibodies Bleeding fevers; Thrombocytopenia (low platelets) Platelet physiology Mega Liver TPO (Thrombopoietin) TPO-receptor No negative feedback to liver Plt Bleeding fevers; Thrombocytopenia (low platelets) Platelet physiology