Roles of TBC1D1 and TBC1D4 in Insulin- and Exercise-Stimulated Glucose Transport of Skeletal Muscle
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

University of California, San Diego
UNIVERSITY OF CALIFORNIA, SAN DIEGO The post-terminal differentiation fate of RNAs revealed by next-generation sequencing A dissertation submitted in partial satisfaction of the requirements for the degree Doctor of Philosophy in Biomedical Sciences by Gloria Kuo Lefkowitz Committee in Charge: Professor Benjamin D. Yu, Chair Professor Richard Gallo Professor Bruce A. Hamilton Professor Miles F. Wilkinson Professor Eugene Yeo 2012 Copyright Gloria Kuo Lefkowitz, 2012 All rights reserved. The Dissertation of Gloria Kuo Lefkowitz is approved, and it is acceptable in quality and form for publication on microfilm and electronically: __________________________________________________________________ __________________________________________________________________ __________________________________________________________________ __________________________________________________________________ __________________________________________________________________ Chair University of California, San Diego 2012 iii DEDICATION Ma and Ba, for your early indulgence and support. Matt and James, for choosing more practical callings. Roy, my love, for patiently sharing the ups and downs of this journey. iv EPIGRAPH It is foolish to tear one's hair in grief, as though sorrow would be made less by baldness. ~Cicero v TABLE OF CONTENTS Signature Page .............................................................................................................. iii Dedication .................................................................................................................... -

Differential Expression Profiling of Gene Response to Ionizing Radiation in Two Endometrial Cancer Cell Lines with Distinct Radiosensitivities
625-634 28/1/2009 12:32 ÌÌ ™ÂÏ›‰·625 ONCOLOGY REPORTS 21: 625-634, 2009 625 Differential expression profiling of gene response to ionizing radiation in two endometrial cancer cell lines with distinct radiosensitivities XUE-LIAN DU1,2, TAO JIANG2, ZE-QING WEN1, QING-SHUI LI2, RONG GAO2 and FEI WANG1 1Department of Gynecologic Oncology, Shandong Tumor Hospital, Shandong University, Jinan 250117; 2Department of Obstetrics and Gynecology, Provincial Hospital Affiliated to Shandong University, Jinan 250021, P.R. China Received November 4, 2008; Accepted December 17, 2008 DOI: 10.3892/or_00000265 Abstracts. Although radiotherapy is routinely administered Introduction to high-risk endometrial carcinoma and offer a significant disease-free survival advantage, the therapeutic effect is Endometrial cancer is one of the most common gynecological sometimes limited by the occurrence of radioresistance. To malignancies worldwide. Surgery is the preferred initial determine the patterns of gene expression responsible for treatment and most women with early-stage, low-risk disease the radioresistance and to search for potential target genes for will do well without adjuvant radiotherapy. However, both radiotherapy, we selected two cell lines with distinct radio- intermediate-risk and high-risk patients are at risk for local- sensitivities using colony-formation assay from four endo- regional relapse and therefore adjuvant radiotherapy, such as metrial cancer cell lines. The cell cycle distribution showed pelvic radiation, vaginal brachytherapy, and whole-abdomen higher fractions of G2/M phase cells in the radiosensitive cell radiation, is essential for local control (1). Johnson and line KLE after radiation compared with the radioresistant cell colleagues reported that adjuvant external-beam pelvic radio- line ISK. -

A Computational Approach for Defining a Signature of Β-Cell Golgi Stress in Diabetes Mellitus
Page 1 of 781 Diabetes A Computational Approach for Defining a Signature of β-Cell Golgi Stress in Diabetes Mellitus Robert N. Bone1,6,7, Olufunmilola Oyebamiji2, Sayali Talware2, Sharmila Selvaraj2, Preethi Krishnan3,6, Farooq Syed1,6,7, Huanmei Wu2, Carmella Evans-Molina 1,3,4,5,6,7,8* Departments of 1Pediatrics, 3Medicine, 4Anatomy, Cell Biology & Physiology, 5Biochemistry & Molecular Biology, the 6Center for Diabetes & Metabolic Diseases, and the 7Herman B. Wells Center for Pediatric Research, Indiana University School of Medicine, Indianapolis, IN 46202; 2Department of BioHealth Informatics, Indiana University-Purdue University Indianapolis, Indianapolis, IN, 46202; 8Roudebush VA Medical Center, Indianapolis, IN 46202. *Corresponding Author(s): Carmella Evans-Molina, MD, PhD ([email protected]) Indiana University School of Medicine, 635 Barnhill Drive, MS 2031A, Indianapolis, IN 46202, Telephone: (317) 274-4145, Fax (317) 274-4107 Running Title: Golgi Stress Response in Diabetes Word Count: 4358 Number of Figures: 6 Keywords: Golgi apparatus stress, Islets, β cell, Type 1 diabetes, Type 2 diabetes 1 Diabetes Publish Ahead of Print, published online August 20, 2020 Diabetes Page 2 of 781 ABSTRACT The Golgi apparatus (GA) is an important site of insulin processing and granule maturation, but whether GA organelle dysfunction and GA stress are present in the diabetic β-cell has not been tested. We utilized an informatics-based approach to develop a transcriptional signature of β-cell GA stress using existing RNA sequencing and microarray datasets generated using human islets from donors with diabetes and islets where type 1(T1D) and type 2 diabetes (T2D) had been modeled ex vivo. To narrow our results to GA-specific genes, we applied a filter set of 1,030 genes accepted as GA associated. -

Supporting Information
Supporting Information Edgar et al. 10.1073/pnas.1601895113 SI Methods (Actimetrics), and recordings were analyzed using LumiCycle Mice. Sample size was determined using the resource equation: Data Analysis software (Actimetrics). E (degrees of freedom in ANOVA) = (total number of exper- – Cell Cycle Analysis of Confluent Cell Monolayers. NIH 3T3, primary imental animals) (number of experimental groups), with −/− sample size adhering to the condition 10 < E < 20. For com- WT, and Bmal1 fibroblasts were sequentially transduced − − parison of MuHV-4 and HSV-1 infection in WT vs. Bmal1 / with lentiviral fluorescent ubiquitin-based cell cycle indicators mice at ZT7 (Fig. 2), the investigator did not know the genotype (FUCCI) mCherry::Cdt1 and amCyan::Geminin reporters (32). of the animals when conducting infections, bioluminescence Dual reporter-positive cells were selected by FACS (Influx Cell imaging, and quantification. For bioluminescence imaging, Sorter; BD Biosciences) and seeded onto 35-mm dishes for mice were injected intraperitoneally with endotoxin-free lucif- subsequent analysis. To confirm that expression of mCherry:: Cdt1 and amCyan::Geminin correspond to G1 (2n DNA con- erin (Promega E6552) using 2 mg total per mouse. Following < ≤ anesthesia with isofluorane, they were scanned with an IVIS tent) and S/G2 (2 n 4 DNA content) cell cycle phases, Lumina (Caliper Life Sciences), 15 min after luciferin admin- respectively, cells were stained with DNA dye DRAQ5 (abcam) and analyzed by flow cytometry (LSR-Fortessa; BD Biosci- istration. Signal intensity was quantified using Living Image ences). To examine dynamics of replicative activity under ex- software (Caliper Life Sciences), obtaining maximum radiance perimental confluent conditions, synchronized FUCCI reporter for designated regions of interest (photons per second per − − − monolayers were observed by time-lapse live cell imaging over square centimeter per Steradian: photons·s 1·cm 2·sr 1), relative 3 d (Nikon Eclipse Ti-E inverted epifluorescent microscope). -

Detecting Novel Effects of Exercise Or AMPK Activation in Human Skeletal Muscle
From the Department of Molecular Medicine and Surgery Karolinska Institutet, Stockholm, Sweden Detecting Novel Effects of Exercise or AMPK Activation in Human Skeletal Muscle David Gray Lassiter Stockholm 2018 All previously published papers were reproduced with permission from the publisher. Published by Karolinska Institutet. Printed by Eprint AB 2018 © David Gray Lassiter, 2018 ISBN 978-91-7831-010-4 Detecting Novel Effects of Exercise or AMPK Activation in Human Skeletal Muscle THESIS FOR DOCTORAL DEGREE (Ph.D.) By David Gray Lassiter Principal Supervisor: Opponent: Juleen Zierath Professor Bret Goodpaster Karolinska Institutet Sanford Burnham Prebys-Medical Discovery Department of Molecular Medicine and Surgery Institute Co-supervisor(s): Examination Board: Anna Krook Professor Anders Arner Karolinska Institutet Karolinska Institutet Department of Physiology and Pharmacology Professor Ewa Ehrenborg Karolinska Institutet Professor Niels Jessen Aarhus University ABSTRACT Cardiovascular and metabolic disorders are among the main causes of death today. Regular exercise can prevent and treat these chronic diseases. A molecule at the center of exercise adaptations in skeletal muscle is adenosine monophosphate-activated protein kinase (AMPK). Rapid energy turnover in cells, such as during contraction in skeletal muscle, activates AMPK. The activation of AMPK leads to inhibition of anabolic processes that consume energy and upregulation of catabolic processes that generate energy. AMPK activation increases glucose uptake into peripheral tissues. Even insulin-resistant individuals, including type 2 diabetes patients, retain the blood-glucose lowering effect of AMPK activation. There is a need to better understand how exercise provides protective benefits, and how AMPK functions at the cellular level. This thesis consists of three research papers wherein exercise and AMPK activation were used as experimental models to identify novel effects of these signals in human skeletal muscle. -

Aneuploidy: Using Genetic Instability to Preserve a Haploid Genome?
Health Science Campus FINAL APPROVAL OF DISSERTATION Doctor of Philosophy in Biomedical Science (Cancer Biology) Aneuploidy: Using genetic instability to preserve a haploid genome? Submitted by: Ramona Ramdath In partial fulfillment of the requirements for the degree of Doctor of Philosophy in Biomedical Science Examination Committee Signature/Date Major Advisor: David Allison, M.D., Ph.D. Academic James Trempe, Ph.D. Advisory Committee: David Giovanucci, Ph.D. Randall Ruch, Ph.D. Ronald Mellgren, Ph.D. Senior Associate Dean College of Graduate Studies Michael S. Bisesi, Ph.D. Date of Defense: April 10, 2009 Aneuploidy: Using genetic instability to preserve a haploid genome? Ramona Ramdath University of Toledo, Health Science Campus 2009 Dedication I dedicate this dissertation to my grandfather who died of lung cancer two years ago, but who always instilled in us the value and importance of education. And to my mom and sister, both of whom have been pillars of support and stimulating conversations. To my sister, Rehanna, especially- I hope this inspires you to achieve all that you want to in life, academically and otherwise. ii Acknowledgements As we go through these academic journeys, there are so many along the way that make an impact not only on our work, but on our lives as well, and I would like to say a heartfelt thank you to all of those people: My Committee members- Dr. James Trempe, Dr. David Giovanucchi, Dr. Ronald Mellgren and Dr. Randall Ruch for their guidance, suggestions, support and confidence in me. My major advisor- Dr. David Allison, for his constructive criticism and positive reinforcement. -
Download Validation Data
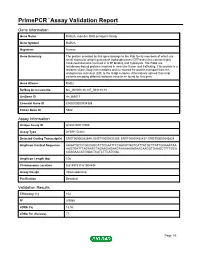
PrimePCR™Assay Validation Report Gene Information Gene Name RAB2A, member RAS oncogene family Gene Symbol RAB2A Organism Human Gene Summary The protein encoded by this gene belongs to the Rab family members of which are small molecular weight guanosine triphosphatases (GTPases) that contain highly conserved domains involved in GTP binding and hydrolysis. The Rabs are membrane-bound proteins involved in vesicular fusion and trafficking. This protein is a resident of pre-Golgi intermediates and is required for protein transport from the endoplasmic reticulum (ER) to the Golgi complex. Alternatively spliced transcript variants encoding different isoforms have been found for this gene. Gene Aliases RAB2 RefSeq Accession No. NC_000008.10, NT_008183.19 UniGene ID Hs.369017 Ensembl Gene ID ENSG00000104388 Entrez Gene ID 5862 Assay Information Unique Assay ID qHsaCID0011980 Assay Type SYBR® Green Detected Coding Transcript(s) ENST00000262646, ENST00000531289, ENST00000452437, ENST00000543829 Amplicon Context Sequence AAGATGCCCGCCAGCATTCCAATTCCAACATGGTCATTATGCTTATTGGAAATAA AAGTGATTTAGAATCTAGAAGAGAAGTAAAAAAAGAAGAAGGTGAAGCTTTTGCA CGAGAACATGGACTCATCTTCATGGA Amplicon Length (bp) 106 Chromosome Location 8:61497317-61504494 Assay Design Intron-spanning Purification Desalted Validation Results Efficiency (%) 104 R2 0.9988 cDNA Cq 18.53 cDNA Tm (Celsius) 77 Page 1/5 PrimePCR™Assay Validation Report gDNA Cq 43.2 Specificity (%) 100 Information to assist with data interpretation is provided at the end of this report. Page 2/5 PrimePCR™Assay Validation Report -

Chemical Denervation Using Botulinum Toxin Increases Akt Expression and Reduces Submaximal Insulin-Stimulated Glucose Transport in Mouse Muscle T
Cellular Signalling 53 (2019) 224–233 Contents lists available at ScienceDirect Cellular Signalling journal homepage: www.elsevier.com/locate/cellsig Chemical denervation using botulinum toxin increases Akt expression and reduces submaximal insulin-stimulated glucose transport in mouse muscle T Zhencheng Lia,1, Lui Näslund-Kocha,b,1, Carlos Henriquez-Olguina, Jonas R. Knudsena, Jingwen Lia, Agnete B. Madsena, Satoru Atod, Jacob Wieneckec, Riki Ogasawarad, ⁎ Jens B. Nielsenb, Thomas E. Jensena, a Section of Molecular Physiology, Department of Nutrition, Exercise and Sports, University of Copenhagen, Denmark b Neural Control of Movement Research Group, Department of Neuroscience, University of Copenhagen, Copenhagen, Denmark c Section of Integrated Physiology, Department of Nutrition, Exercise and Sports, University of Copenhagen, Denmark d Nagoya Institute of Technology, Nagoya, Japan ARTICLE INFO ABSTRACT Keywords: Botulinum toxin A (botox) is a toxin used for spasticity treatment and cosmetic purposes. Botox blocks the Atrophy excitation of skeletal muscle fibers by preventing the release of acetylcholine from motor nerves, a process GLUT4 termed chemical denervation. Surgical denervation is associated with increased expression of the canonical Hexokinase insulin-activated kinase Akt, lower expression of glucose handling proteins GLUT4 and hexokinase II (HKII) and TBC1D4 insulin resistant glucose uptake, but it is not known if botox has a similar effect. To test this, we performed a Insulin signaling time-course study using supra-maximal insulin-stimulation in mouse soleus ex vivo. No effect was observed in the glucose transport responsiveness at day 1, 7 and 21 after intramuscular botox injection, despite lower ex- pression of GLUT4, HKII and expression and phosphorylation of TBC1D4. -

Bioinformatics Analysis to Screen Key Genes in Papillary Thyroid Carcinoma
ONCOLOGY LETTERS 19: 195-204, 2020 Bioinformatics analysis to screen key genes in papillary thyroid carcinoma YUANHU LIU1*, SHUWEI GAO2*, YAQIONG JIN2, YERAN YANG2, JUN TAI1, SHENGCAI WANG1, HUI YANG2, PING CHU2, SHUJING HAN2, JIE LU2, XIN NI1,2, YONGBO YU2 and YONGLI GUO2 1Department of Otolaryngology, Head and Neck Surgery, Beijing Children's Hospital, Capital Medical University, National Center for Children's Health; 2Beijing Key Laboratory for Pediatric Diseases of Otolaryngology, Head and Neck Surgery, MOE Key Laboratory of Major Diseases in Children, Beijing Pediatric Research Institute, Beijing Children's Hospital, Capital Medical University, National Center for Children's Health, Beijing 100045, P.R. China Received April 22, 2019; Accepted September 24, 2019 DOI: 10.3892/ol.2019.11100 Abstract. Papillary thyroid carcinoma (PTC) is the most verifying their potential for clinical diagnosis. Taken together, common type of thyroid carcinoma, and its incidence has the findings of the present study suggest that these genes and been on the increase in recent years. However, the molecular related pathways are involved in key events of PTC progression mechanism of PTC is unclear and misdiagnosis remains a and facilitate the identification of prognostic biomarkers. major issue. Therefore, the present study aimed to investigate this mechanism, and to identify key prognostic biomarkers. Introduction Integrated analysis was used to explore differentially expressed genes (DEGs) between PTC and healthy thyroid tissue. To Thyroid carcinoma is the most common malignancy of investigate the functions and pathways associated with DEGs, the head and neck, and accounts for 91.5% of all endocrine Gene Ontology, pathway and protein-protein interaction (PPI) malignancies (1). -

The Rabgaps TBC1D1 and TBC1D4 Control Uptake of Long-Chain Fatty
Diabetes Page 2 of 39 1 The RabGAPs TBC1D1 and TBC1D4 control uptake of long-chain fatty 2 acids into skeletal muscle via fatty acid transporter SLC27A4/FATP4 3 Short title: RabGAPs in skeletal muscle lipid metabolism 4 Tim Benninghoff1,2, Lena Espelage1,2, Samaneh Eickelschulte1,2, Isabel Zeinert1, Isabelle 5 Sinowenka1, Frank Müller1, Christina Schöndeling1, Hannah Batchelor1, Sandra Cames1,2, 6 Zhou Zhou1, Jörg Kotzka1,2, Alexandra Chadt §1,2 and Hadi Al-Hasani1,2 7 8 1Institute for Clinical Biochemistry and Pathobiochemistry, German Diabetes Center, Leibniz 9 Center for Diabetes Research at Heinrich Heine University Duesseldorf 10 2German Center for Diabetes Research, München-Neuherberg, Germany. 11 12 § Corresponding author 13 Dr. Alexandra Chadt 14 Institute for Clinical Biochemistry and Pathobiochemistry 15 German Diabetes Center, Leibniz Center for Diabetes Research at Heinrich Heine University 16 Duesseldorf 17 Auf'm Hennekamp 65 18 40225 Duesseldorf 19 Germany 20 tel/fax +49-211-3382-577/ -430 21 [email protected] 22 23 Word count: 4946 (Introduction, Methods, Results, Discussion) 24 Figures: 5 25 Tables: 0 1 Diabetes Publish Ahead of Print, published online August 31, 2020 Page 3 of 39 Diabetes 26 Abstract 27 The two closely related RabGTPase-activating proteins (RabGAPs) TBC1D1 and TBC1D4 28 play a crucial role in the regulation of GLUT4 translocation in response to insulin and 29 contraction in skeletal muscle. In mice, deficiency in one or both RabGAPs leads to reduced 30 insulin and contraction-stimulated glucose uptake, and to elevated fatty acid uptake and 31 oxidation in both glycolytic and oxidative muscle fibers without altering mitochondrial copy 32 number and the abundance of OXPHOS proteins. -

Prior AICAR Stimulation Increases Insulin Sensitivity in Mouse Skeletal Muscle in an AMPK-Dependent Manner
2042 Diabetes Volume 64, June 2015 Rasmus Kjøbsted,1,2 Jonas T. Treebak,1,2 Joachim Fentz,1 Louise Lantier,3,4,5 Benoit Viollet,3,4,5 Jesper B. Birk,1 Peter Schjerling,6 Marie Björnholm,7 Juleen R. Zierath,2,7 and Jørgen F.P. Wojtaszewski1 Prior AICAR Stimulation Increases Insulin Sensitivity in Mouse Skeletal Muscle in an AMPK-Dependent Manner Diabetes 2015;64:2042–2055 | DOI: 10.2337/db14-1402 An acute bout of exercise increases glucose uptake in rodents (1–6) and may persist for up to 48 h after exercise, skeletal muscle by an insulin-independent mechanism. depending on carbohydrate availability (7–9). Improved mus- In the period after exercise, insulin sensitivity to in- cle insulin sensitivity postexercise is mediated by one or creased glucose uptake is enhanced. The molecular several local contraction-induced mechanisms (10) involving mechanisms underpinning this phenomenon are poorly both enhanced transport and intracellular processing of glu- understood but appear to involve an increased cell cose. This period is characterized by increased GLUT4 protein surface abundance of GLUT4. While increased proximal abundance at the plasma membraneandenhancedglycogen insulin signaling does not seem to mediate this effect, synthase activity (11,12). These changes occur independent elevated phosphorylation of TBC1D4, a downstream target of both insulin (Akt) and exercise (AMPK) signal- of global protein synthesis (13), including both total GLUT4 ing, appears to play a role. The main purpose of this and glycogen synthase protein content (4,11), and are inde- study was to determine whether AMPK activation pendent of changes in proximal insulin signaling, including increases skeletal muscle insulin sensitivity. -

A SARS-Cov-2-Human Protein-Protein Interaction Map Reveals Drug Targets and Potential Drug- Repurposing
bioRxiv preprint doi: https://doi.org/10.1101/2020.03.22.002386; this version posted March 23, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY 4.0 International license. A SARS-CoV-2-Human Protein-Protein Interaction Map Reveals Drug Targets and Potential Drug- Repurposing David E. Gordon1,2,3,4, Gwendolyn M. Jang1,2,3,4, Mehdi Bouhaddou1,2,3,4, Jiewei Xu1,2,3,4, Kirsten Obernier1,2,3,4, Matthew J. O'Meara5, Jeffrey Z. Guo1,2,3,4, Danielle L. Swaney1,2,3,4, Tia A. Tummino1,2,6, Ruth Huettenhain1,2,3,4, Robyn Kaake1,2,3,4, Alicia L. Richards1,2,3,4, Beril Tutuncuoglu1,2,3,4, Helene Foussard1,2,3,4, Jyoti Batra1,2,3,4, Kelsey Haas1,2,3,4, Maya Modak1,2,3,4, Minkyu Kim1,2,3,4, Paige Haas1,2,3,4, Benjamin J. Polacco1,2,3,4, Hannes Braberg1,2,3,4, Jacqueline M. Fabius1,2,3,4, Manon Eckhardt1,2,3,4, Margaret Soucheray1,2,3,4, Melanie J. Bennett1,2,3,4, Merve Cakir1,2,3,4, Michael J. McGregor1,2,3,4, Qiongyu Li1,2,3,4, Zun Zar Chi Naing1,2,3,4, Yuan Zhou1,2,3,4, Shiming Peng1,2,6, Ilsa T. Kirby1,4,7, James E. Melnyk1,4,7, John S. Chorba1,4,7, Kevin Lou1,4,7, ShiZhong A. Dai1,4,7, Wenqi Shen1,4,7, Ying Shi1,4,7, Ziyang Zhang1,4,7, Inigo Barrio-HernandeZ8, Danish Memon8, Claudia Hernandez-Armenta8, Christopher J.P.