Engrailed-Like Immunoreactivity in the Embryonic Ventral Nerve Cord of the Marbled Crayfish
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

T-Box Genes in Limb Development and Disease
Open Research Online The Open University’s repository of research publications and other research outputs T-box Genes in Limb Development and Disease Thesis How to cite: Rallis, Charalampos (2004). T-box Genes in Limb Development and Disease. PhD thesis The Open University. For guidance on citations see FAQs. c 2004 Charalampos Rallis Version: Version of Record Link(s) to article on publisher’s website: http://dx.doi.org/doi:10.21954/ou.ro.0000fa0b Copyright and Moral Rights for the articles on this site are retained by the individual authors and/or other copyright owners. For more information on Open Research Online’s data policy on reuse of materials please consult the policies page. oro.open.ac.uk T-box Genes in Limb Development and Disease Charalampos Rallis Thesis submitted for the degree of Doctor of Philosophy October 2004 Division of Developmental Biology National Institute for Medical Research Mill Hill London Open University ProQuest Number: C819643 All rights reserved INFORMATION TO ALL USERS The quality of this reproduction is dependent upon the quality of the copy submitted. In the unlikely event that the author did not send a com plete manuscript and there are missing pages, these will be noted. Also, if material had to be removed, a note will indicate the deletion. uest ProQuest C819643 Published by ProQuest LLO (2019). Copyright of the Dissertation is held by the Author. All rights reserved. This work is protected against unauthorized copying under Title 17, United States C ode Microform Edition © ProQuest LLO. ProQuest LLO. 789 East Eisenhower Parkway P.Q. -

Pancreas-Specific Deletion of Mouse Gata4 and Gata6 Causes Pancreatic Agenesis
Pancreas-specific deletion of mouse Gata4 and Gata6 causes pancreatic agenesis Shouhong Xuan, … , Raymond J. Macdonald, Lori Sussel J Clin Invest. 2012;122(10):3516-3528. https://doi.org/10.1172/JCI63352. Research Article Pancreatic agenesis is a human disorder caused by defects in pancreas development. To date, only a few genes have been linked to pancreatic agenesis in humans, with mutations in pancreatic and duodenal homeobox 1 (PDX1) and pancreas-specific transcription factor 1a (PTF1A) reported in only 5 families with described cases. Recently, mutations in GATA6 have been identified in a large percentage of human cases, and aG ATA4 mutant allele has been implicated in a single case. In the mouse, Gata4 and Gata6 are expressed in several endoderm-derived tissues, including the pancreas. To analyze the functions of GATA4 and/or GATA6 during mouse pancreatic development, we generated pancreas- specific deletions of Gata4 and Gata6. Surprisingly, loss of either Gata4 or Gata6 in the pancreas resulted in only mild pancreatic defects, which resolved postnatally. However, simultaneous deletion of both Gata4 and Gata6 in the pancreas caused severe pancreatic agenesis due to disruption of pancreatic progenitor cell proliferation, defects in branching morphogenesis, and a subsequent failure to induce the differentiation of progenitor cells expressing carboxypeptidase A1 (CPA1) and neurogenin 3 (NEUROG3). These studies address the conserved and nonconserved mechanisms underlying GATA4 and GATA6 function during pancreas development and provide a new mouse model to characterize the underlying developmental defects associated with pancreatic agenesis. Find the latest version: https://jci.me/63352/pdf Research article Related Commentary, page 3469 Pancreas-specific deletion of mouse Gata4 and Gata6 causes pancreatic agenesis Shouhong Xuan,1 Matthew J. -

Homologues of the Engrailed Gene from Five Molluscan Classes
FEBS 15444 FEBS Letters 365 (1995) 71 74 Homologues of the engrailed gene from five molluscan classes C.G. Wray a, D.K. Jacobs a'*, R. Kostriken b, A.P. Vogler c, R. Baker d, R. DeSalle d "Department of Biology, University of California, 621 Circle Drive South, Los Angeles, CA 90095-1606, USA bDepartment of Cell Biology and Anatomy, New York Medical College, Valhalla, NY 10595, USA ~Department of Entomology, The Natural History Museum, Cromwell Road, London, SW7 5BD, UK JAmerican Museum of Natural History, Central Park West at 79th Street, New York, NY 10024, USA Received 1 March 1995 potential homology and phylogenetic significance of metamer- Abstract We used the polymerase chain reaction (PCR) to am- ism amongst the protostomes. plify, clone, and sequence 10 engrailed homeodomains from Here we report DNA sequence of the engrailed gene from 8 species in the five major molluscan classes, including the serially eight molluscs representing the five major classes in the phylum. organized chiton (Polyplacophora) lineage. The Drosophila melanogaster gene engrailed (en) is one of several genes involved A 232 base pair segment including the engrailed homeodomain in embyonic segment polarity determination. Studies of engraUed and an engrailed specific region 3' to the homeodomain was sequence and expression in molluscs are of interest due to ques- amplified using the polymerase chain reaction (PCR), cloned tions regarding the evolution and homology of segmentation in and subsequently sequenced. these taxa. Nucleotide and deduced amino acid sequence compar- isons reflect evolutionary conservation within helices of the en 2. Materials and methods homeodomain and ancient divergences in the region 3' to the homeodomain. -

Influence of Habitat and Bat Activity on Moth Community Composition and Seasonal Phenology Across Habitat Types
INFLUENCE OF HABITAT AND BAT ACTIVITY ON MOTH COMMUNITY COMPOSITION AND SEASONAL PHENOLOGY ACROSS HABITAT TYPES BY MATTHEW SAFFORD THESIS Submitted in partial fulfillment of the requirements for the degree of Master of Science in Entomology in the Graduate College of the University of Illinois at Urbana-Champaign, 2018 Urbana, Illinois Advisor: Assistant Professor Alexandra Harmon-Threatt, Chair and Director of Research ABSTRACT Understanding the factors that influence moth diversity and abundance is important for monitoring moth biodiversity and developing conservation strategies. Studies of moth habitat use have primarily focused on access to host plants used by specific moth species. How vegetation structure influences moth communities within and between habitats and mediates the activity of insectivorous bats is understudied. Previous research into the impact of bat activity on moths has primarily focused on interactions in a single habitat type or a single moth species of interest, leaving a large knowledge gap on how habitat structure and bat activity influence the composition of moth communities across habitat types. I conducted monthly surveys at sites in two habitat types, restoration prairie and forest. Moths were collected using black light bucket traps and identified to species. Bat echolocation calls were recorded using ultrasonic detectors and classified into phonic groups to understand how moth community responds to the presence of these predators. Plant diversity and habitat structure variables, including tree diameter at breast height, ground cover, and vegetation height were measured during summer surveys to document how differences in habitat structure between and within habitats influences moth diversity. I found that moth communities vary significantly between habitat types. -

Sequential Organizing Activities of Engrailed, Hedgehog and Decapentaplegic in the Drosophila Wing
Development 121, 2265-2278 (1995) 2265 Printed in Great Britain © The Company of Biologists Limited 1995 Sequential organizing activities of engrailed, hedgehog and decapentaplegic in the Drosophila wing Myriam Zecca1, Konrad Basler1 and Gary Struhl2 1Zoologisches Institut, Universität Zürich, Winterthurerstrasse 190, 8057 Zürich, Switzerland 2Howard Hughes Medical Institute, Department of Genetics and Development, Columbia University College of Physicians and Surgeons, 701 West 168th Street, New York NY 10032 USA SUMMARY The Drosophila wing is formed by two cell populations, the strate that dpp can exert a long-range organizing influence anterior and posterior compartments, which are distin- on surrounding wing tissue, specifying anterior or posterior guished by the activity of the selector gene engrailed (en) in pattern depending on the compartmental provenance, and posterior cells. Here, we show that en governs growth and hence the state of en activity, of the responding cells. Thus, patterning in both compartments by controlling the dpp secreted by anterior cells along the compartment expression of the secreted proteins hedgehog (hh) and boundary has the capacity to organize the development of decapentaplegic (dpp) as well as the response of cells to both compartments. Finally, we report evidence suggesting these signaling molecules. First, we demonstrate that en that dpp may exert its organizing influence by acting as a activity programs wing cells to express hh whereas the gradient morphogen in contrast to hh which appears to act absence of en activity programs them to respond to hh by principally as a short range inducer of dpp. expressing dpp. As a consequence, posterior cells secrete hh and induce a stripe of neighboring anterior cells across the Key words: engrailed, decapentaplegic, hedgehog, Drosophila, compartment boundary to secrete dpp. -

Toby Austin's Garden Moth List
Toby Austin’s Garden Moth List Orange Swift Triodia sylvina Common Swift Korscheltellus lupulinus Ghost Moth Hepialus humuli Nematopogon swammerdamella Cork Moth Nemapogon cloacella Tinea trinotella Horse-chestnut Leaf-miner Cameraria ohridella Bird-cherry Ermine Yponomeuta evonymella Orchard/Apple/Spindle Ermine Yponomeuta padella/malinellus/cagnagella Willow Ermine Yponomeuta rorrella Yponomeuta plumbella Ypsolopha ustella Diamond-back Moth Plutella xylostella Plutella porrectella Ash Bud Moth Prays fraxinella Hawthorn Moth Scythropia crataegella Oegoconia quadripuncta/caradjai/deauratella Crassa unitella Carcina quercana Luquetia lobella Agonopterix alstromeriana Mompha sturnipennella Blastobasis adustella Blastobasis lacticolella Twenty-plume Moth Alucita hexadactyla Beautiful Plume Amblyptilia acanthadactyla Stenoptilia pterodactyla Stenoptilia bipunctidactyla Common Plume Emmelina monodactyla Variegated Golden Tortrix Archips xylosteana Argyrotaenia ljungiana Chequered Fruit-tree Tortrix Pandemis corylana Dark Fruit-tree Tortrix Pandemis heparana Pandemis dumetana Syndemis musculana Lozotaenia forsterana Carnation Tortrix Cacoecimorpha pronubana Light Brown Apple Moth Epiphyas postvittana Lozotaeniodes formosana Summer Fruit Tortrix Adoxophyes orana Flax Tortrix Cnephasia asseclana Green Oak Tortrix Tortrix viridana Cochylis molliculana Cochylis atricapitana Acleris holmiana Acleris forsskaleana Acleris comariana/laterana Acleris cristana Garden Rose Tortrix Acleris variegana Pseudargyrotoza conwagana Phtheochroa rugosana Agapeta -
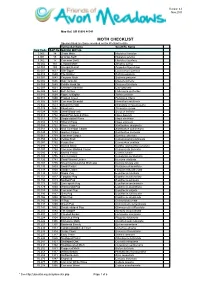
MOTH CHECKLIST Species Listed Are Those Recorded on the Wetland to Date
Version 4.0 Nov 2015 Map Ref: SO 95086 46541 MOTH CHECKLIST Species listed are those recorded on the Wetland to date. Vernacular Name Scientific Name New Code B&F No. MACRO MOTHS 3.005 14 Ghost Moth Hepialus humulae 3.001 15 Orange Swift Hepialus sylvina 3.002 17 Common Swift Hepialus lupulinus 50.002 161 Leopard Moth Zeuzera pyrina 54.008 169 Six-spot Burnet Zygaeba filipendulae 66.007 1637 Oak Eggar Lasiocampa quercus 66.010 1640 The Drinker Euthrix potatoria 68.001 1643 Emperor Moth Saturnia pavonia 65.002 1646 Oak Hook-tip Drepana binaria 65.005 1648 Pebble Hook-tip Drepana falcataria 65.007 1651 Chinese Character Cilix glaucata 65.009 1653 Buff Arches Habrosyne pyritoides 65.010 1654 Figure of Eighty Tethia ocularis 65.015 1660 Frosted Green Polyploca ridens 70.305 1669 Common Emerald Hermithea aestivaria 70.302 1673 Small Emerald Hemistola chrysoprasaria 70.029 1682 Blood-vein Timandra comae 70.024 1690 Small Blood-vein Scopula imitaria 70.013 1702 Small Fan-footed Wave Idaea biselata 70.011 1708 Single-dotted Wave Idaea dimidiata 70.016 1713 Riband Wave Idaea aversata 70.053 1722 Flame Carpet Xanthorhoe designata 70.051 1724 Red Twin-spot Carpet Xanthorhoe spadicearia 70.049 1728 Garden Carpet Xanthorhoe fluctuata 70.061 1738 Common Carpet Epirrhoe alternata 70.059 1742 Yellow Shell Camptogramma bilineata 70.087 1752 Purple Bar Cosmorhoe ocellata 70.093 1758 Barred Straw Eulithis (Gandaritis) pyraliata 70.097 1764 Common Marbled Carpet Chloroclysta truncata 70.085 1765 Barred Yellow Cidaria fulvata 70.100 1776 Green Carpet Colostygia pectinataria 70.126 1781 Small Waved Umber Horisme vitalbata 70.107 1795 November/Autumnal Moth agg Epirrita dilutata agg. -

Heraldry in Ireland
Heraldry in Ireland Celebrating 75 years of the Office of the Chief Herald at the NLI Sir John Ainsworth Shield Vert, a chevron between three battle-axes argent Crest A falcon rising proper, beaked, legged and belled gules Motto Surgo et resurgam Did you know? Sir John Ainsworth was the NLI's Surveyor of Records in Private Keeping in the 1940s and 1950s. Roderick More OFerrall Shield Quarterly: 1st, Vert, a lion rampant or (for O Ferrall); 2nd, Vert a lion rampant in chief three estoiles or (for O More); 3rd, Argent, upon a mount vert two lions rampant combatant gules supporting the trunk of an oak tree entwined with a serpent descending proper, (for O Reilly); 4th, Azure, a bend cotised or between six escallops argent (for Cruise) Crest On a ducal coronet or a greyhound springing sable; A dexter hand lying fess-ways proper cuffed or holding a sword in pale hilted of the second pierced through three gory heads of the first Motto Cú re bu; Spes mea Deus Did you know? This four designs on the shield represent four families. Heiress Leticia More of Balyna, county Kildare married Richard Ferrall in 1751. Their grandson Charles Edward More O'Ferrall married Susan O'Reilly in 1849. Susan was the daughter of Dominic O'Reilly of Kildangan Castle, county Kildare who had married heiress Susanna Cruise in 1818. Dublin Stock Exchange Shield Quarterly: 1st, Sable, a tower or; 2nd, Vert, three swords points upwards two and one proper pommelled and hilted or; 3rd, Vert, three anchors erect two and one argent; 4th, Chequy, sable and argent, on a chief argent an escroll proper, inscribed thereon the words Geo. -

Quelques Papillons De Nuit De La Réserve Faunique De Matane
Quelques papillons de nuit de la réserve faunique de Matane Le mont Blanc à l’arrière-plan Comité de protection des monts Chic-Chocs Rapport produit par Jacques Larivée Rimouski, mai 2017 Papillons de nuit de la réserve faunique de Matane C’est à l’invitation du Comité de protection des monts Chic-Chocs que je joins du 15 au 17 août 2016 un groupe de naturalistes qui a pour objectif d’améliorer les connaissances de la flore et de la faune du territoire de la réserve faunique de Matane. Voici les membres de l’équipe dans l’ordre où ils apparaissent sur la photo. Photo Claude Gauthier Claude Gauthier (ornithologie, photographie, kayak, transport et sécurité) ; Christian Grenier (botanique des plantes vasculaires et photographie) ; Pierre Fradette (organisation, ornithologie, photographie, transport et sécurité), Louis Fradette (organisation, ornithologie, photographie, kayak, transport et sécurité) ; Pierre Lévesque (bryologie et photographie) ; Jean Faubert (bryologie et photographie) ; Jacques Larivée (ornithologie, photographie et entomologie) ; Gaétan Caron (organisation, transport, connaissance du territoire et sécurité). 2 Papillons de nuit de la réserve faunique de Matane Mon « travail » consiste à noter mes observations des oiseaux le jour partout sur le territoire, comme le font les autres ornithologues, et à photographier les papillons de nuit le soir à l’Étang à la Truite le 15 et le 17 août et au sommet du mont Blanc le soir du 16 août. Mes observations d’oiseaux sont enregistrées sur eBird et sont résumées dans ce document. Le texte qui suit présente 3 listes d’espèces incluant au moins une photo par espèce : la liste des papillons de nuit photographiés à l’Étang à la Truite suivie de la liste des papillons de nuit photographiés au mont Blanc et de la liste des papillons de nuit non identifiés. -

Engrailed Homeoproteins in Visual System Development
Cell. Mol. Life Sci. (2015) 72:1433–1445 DOI 10.1007/s00018-014-1776-z Cellular and Molecular Life Sciences REVIEW Engrailed homeoproteins in visual system development Andrea Wizenmann • Olivier Stettler • Kenneth L. Moya Received: 28 July 2014 / Revised: 31 October 2014 / Accepted: 6 November 2014 / Published online: 29 November 2014 Ó The Author(s) 2014. This article is published with open access at Springerlink.com Abstract Engrailed is a homeoprotein transcription Keywords Visual system Á Retina Á Tectum Á factor. This family of transcription factors is characterized Sensory map Á Homeoprotein Á Engrailed by their DNA-binding homeodomain and some members, including Engrailed, can transfer between cells and reg- ulate protein translation in addition to gene transcription. Introduction Engrailed is intimately involved in the development of the vertebrate visual system. Early expression of Engrailed in Since the discovery of homeobox genes [1, 2] there has dorsal mesencephalon contributes to the development and been accumulating evidence from all multi-cellular organization of a visual structure, the optic tectum/supe- organisms that these genes play key roles in determining rior colliculus. This structure is an important target for positional information. These genes encode homeoprotein retinal ganglion cell axons that carry visual information transcription factors that regulate the expression of down- from the retina. Engrailed regulates the expression of stream genes necessary at all developmental stages, Ephrin axon guidance cues in the tectum/superior col- including lineage determination, cell migration, cell dif- liculus. More recently it has been reported that Engrailed ferentiation, and tissue formation. Some homeoproteins are itself acts as an axon guidance cue in synergy with the also able to regulate protein translation and cell-to-cell Ephrin system and is proposed to enhance retinal topo- signaling. -

Transcriptional and Epigenetic Control of Brown and Beige Adipose Cell Fate and Function
REVIEWS Transcriptional and epigenetic control of brown and beige adipose cell fate and function Takeshi Inagaki1,2, Juro Sakai1,2 and Shingo Kajimura3 Abstract | White adipocytes store excess energy in the form of triglycerides, whereas brown and beige adipocytes dissipate energy in the form of heat. This thermogenic function relies on the activation of brown and beige adipocyte-specific gene programmes that are coordinately regulated by adipose-selective chromatin architectures and by a set of unique transcriptional and epigenetic regulators. A number of transcriptional and epigenetic regulators are also required for promoting beige adipocyte biogenesis in response to various environmental stimuli. A better understanding of the molecular mechanisms governing the generation and function of brown and beige adipocytes is necessary to allow us to control adipose cell fate and stimulate thermogenesis. This may provide a therapeutic approach for the treatment of obesity and obesity-associated diseases, such as type 2 diabetes. Interscapular BAT Adipose tissue has a central role in whole-body energy subjects who had previously lacked detectable BAT Brown adipose tissue (BAT) is a homeostasis. White adipose tissue (WAT) is the major depots before cold exposure, presumably owing to the specialized organ that adipose organ in mammals. It represents 10% or more emergence of new thermogenic adipocytes. This, then, produces heat. BAT is localized of the body weight of healthy adult humans and is leads to an increase in non-shivering thermogenesis in the interscapular and 6–9 perirenal regions of rodents specialized for the storage of excess energy. Humans and/or an improvement in insulin sensitivity . These and infants. -

Beginner S Guide to Moths of the Midwest Geometers
0LGZHVW5HJLRQ86$ %HJLQQHU V*XLGHWR0RWKVRIWKH0LGZHVW*HRPHWHUV $QJHOOD0RRUHKRXVH ,OOLQRLV1DWXUH3UHVHUYH&RPPLVVLRQ Photos: Angella Moorehouse ([email protected]). Produced by: Angella Moorehouse with the assistance of Alicia Diaz, Field Museum. Identification assistance provided by: multiple sources (inaturalist.org; bugguide.net) )LHOG0XVHXP &&%<1&/LFHQVHGZRUNVDUHIUHHWRXVHVKDUHUHPL[ZLWKDWWULEXWLRQEXWFRPPHUFLDOXVHRIWKHRULJLQDOZRUN LVQRWSHUPLWWHG >ILHOGJXLGHVILHOGPXVHXPRUJ@>@YHUVLRQ $ERXWWKH%(*,11(5¶6027+62)7+(0,':(67*8,'(6 Most photos were taken in west-central and central Illinois; a fewDUH from eastern Iowa and north-central Wisconsin. Nearly all were posted to identification websites: BugGuide.netDQG iNaturalist.org. Identification help was provided by Aaron Hunt, Steve Nanz, John and Jane Balaban, Chris Grinter, Frank Hitchell, Jason Dombroskie, William H. Taft, Jim Wiker,DQGTerry Harrison as well as others contributing to the websites. Attempts were made to obtain expert verifications for all photos to the field identification level, however, there will be errors. Please contact the author with all corrections Additional assistance was provided by longtime Lepidoptera survey partner, Susan Hargrove. The intention of these guides is to provide the means to compare photographs of living specimens of related moths from the Midwest to aid the citizen scientists with identification in the field for Bio Blitz, Moth-ers Day, and other night lighting events. A taxonomic list to all the species featured is provided at the end along with some field identification tips. :(%6,7(63529,',1*,'(17,),&$7,21,1)250$7,21 BugGuide.net LNaturalist.org Mothphotographersgroup.msstate.edu Insectsofiowa.org centralillinoisinsects.org/weblog/resources/ :+,&+027+*8,'(7286( The moths were split into 6 groups for the purposes of creating smaller guides focusing on similar features of 1 or more superfamilies.