I the FLAVONOID APIGENIN INHIBITS INDUCIBLE
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Estroquench™ Hormone Specific Formulation™
PRODUCT DATA DOUGLAS LABORATORIES® 08/2014 1 EstroQuench™ Hormone Specific Formulation™ DESCRIPTION EstroQuench™ is a Hormone Specific Formulation™ of ingredients that have documented anti-aromatase activity as well as androgenic adaptogens which support the function of endogenous aromatase inhibitors. Collectively these herbs promote minimal production and function of estrogens, while promoting testosterone function, including optimal sexual function in both genders. This formulation is designed to quench excessive production of estrogens and aberrant functions of them while supporting optimal function of androgens by maintaining the health of androgen producing glands.† Hormone Specific Formulation™ provided by Douglas Laboratories® and formulated by Dr. Joseph J Collins is created to support the optimal function of specific hormones through the use of hormone specific adaptogens, hormone specific agonists and hormone specific functional mimetics. This formulation may be used as part of a hormone health program with dietary and nutrient support. In addition, this formulation may be used by clinicians as an adjuvant to support optimal hormone health in patients who have been prescribed bioidentical hormone therapies. FUNCTIONS Aromatase (a cytochrome P450 enzyme {CYP19}) is the enzyme that controls the conversion of androgens to estrogens. More specifically, aromatase is the enzyme responsible for catalyzing the biosynthesis of androstenedione into estrone, and the biosynthesis of testosterone to estradiol. Estrogens include the broad range of aromatized hormones created form androgens. The specific attribute of estrogens that separate them from progestogens, androgens and corticoids is that estrogens are the only aromatized steroid hormones. Estradiol, the most potent endogenous estrogen, is biosynthesized by aromatization from androgens by aromatase (which is also called estrogen synthase). -

Test Report Comprehensive Hormone Insights™
698814 COMPREHENSIVE HORMONE INSIGHTS™ TEST REPORT Dr. Maximus, N.D. E: [email protected] Date of Collection: P: 403-241-4500 Time of Collection: F: 403-241-4501 Date of Receipt: www.rmalab.com Reported On: CHI Accession: 698814 Healthcare Professional Patient Age: Dr. Maximus, N.D. Date of Birth: Gender: Male F: Relevant Medications Biometrics Curcumin Height (in) : 73 Weight (lb) : 180 BMI : 24 Waist (in) : 35 Hip (in) : 41 CHI Accession: 698814 SUMMARY HMUS01 How to read the graphs LEGEND: 50 66 Sex Steroid Hormones 50 66 Middle third of 33 33 84 reference population Hormone Start of 83 100 80 100 highest 16 Percentile Precursors 16 Percentile third of Sum of Androgens Sum of Estrogens 50 66 reference 0 0 population (T, DHT, α+β androstanediol) Listed in Interp Guide 33 84 End of 100 16 lowest Percentile00 50 66 50 66 third of 33 33 84 reference 0 population Patient’s percentile rank 81 100 95 100 compared to reference 16 Percentile 16 Percentile population (see summary) DHEA + Metabolites Sum of Progesterone Metabolites 0 (DHEA + A + E) 0 α+β Pregnanediol Cortisol Melatonin Oxidative Stress Free Cortisol Profile (ng/mg) 100 50 66 50 66 33 84 33 84 80 64 100 0 100 16 Percentile 16 Percentile 60 6-sulfatoxy 8-Hydroxy-2- 0 Melatonin 0 deoxyguanosine 40 (Overnight) (Overnight) 20 6-sulfatoxymelatonin provides 8-hydroxy-2-deoxyguanosine is Cortisol/Creatinine (ng/mg) insight into melatonin levels. a marker of oxidative stress 0 Morning Dinner Bedtime A B C 50 66 Free cortisol Cortisol Metabolites 33 84 profile is used to provides a general Testosterone Cortisol assess diurnal assessment of 16 100 cortisol rhythm adrenal cortisol 16 Percentile Cortisol production Cortisol Metabolites 0 (α+β THF + THE) Testosterone Cortisol/Testosterone provides insight into relative catabolic (cortisol) and anabolic (testosterone) states. -

2018 Apigenin Male Infertility (2).Pdf
Original Article Thai Journal of Pharmaceutical Sciences Apigenin and baicalin, each alone or in low-dose combination, attenuated chloroquine induced male infertility in adult rats Amira Akilah, Mohamed Balaha*, Mohamed-Nabeih Abd-El Rahman, Sabiha Hedya Department of Pharmacology, Faculty of Medicine, Tanta University, Postal No. 31527, El-Gish Street, Tanta, Egypt Corresponding Author: Department of Pharmacology, ABSTRACT Faculty of Medicine, Tanta University, Postal No. 31527, Introduction: Male infertility is a worldwide health problem, which accounts for about 50% of all El-Gish Street, Tanta, Egypt. cases of infertility and considered as the most common single defined cause of infertility. Recently, Tel.: +201284451952. E-mail: apigenin and baicalin exhibited a powerful antioxidant and antiapoptotic activities. Consequently, in [email protected]. the present study, we evaluated the possible protective effect of apigenin and baicalin, either alone or Edu.Eg (M. Balaha). in low-dose combination, on a rat model of male infertility, regarding for its effects on the hormonal assay, testicular weight, sperm parameters, oxidative-stress state, apoptosis, and histopathological Received: Mar 06, 2018 changes. Material and methods: 12-week-old adult male Wister rats received 10 mg/kg/d Accepted: May 08, 2018 chloroquine orally for 30 days to induce male infertility. Either apigenin (30 or 15 mg/kg/d), baicalin Published: July 10, 2018 (100 or 50 mg/kg/d) or a combination of 15 mg/kg/d apigenin and 50 mg/kg/d baicalin received daily -

(Apigenin, Luteolin, Chrysin) and Their Importance for Health
Mellifera 2020, 20(1):16-27 MELLIFERA REVIEW ARTICLE Flavones (Apigenin, Luteolin, Chrysin) and Their Importance for Health Fatma Özsel ÖZCAN1*, Ozan ALDEMİR2, Bülent KARABULUT3 1Beykoz Üniversity, Faculty of Art and Design, Department of Gastronomy and Culinary Arts, İstanbul, Turkey 2Ege University Faculty of Medicine, Department of Biochemistry, İzmir, TURKEY 3Ege University Faculty of Medicine, Department of Oncology, İzmir, TURKEY * Corresponding author e-mail: [email protected] Received: 14th March, 2020; accepted: 4th May, 2020; published: 29th June, 2020 A B S T R A C T It has been shown in recent years that foods called functional foods may protect against certain types of cancer, cardiovascular diseases and cognitive dysfunctions. In the studies performed, the flavonoids (apigenin, chrysin, luteolin) which are subclass of flavonoids have been shown to have antioxidant, antiinflammatory, antiallergic, neuroprotective and cardioprotective effects and it is presented as the current treatment method in the treatment of some diseases. The structure function, nutritional resources and potential therapeutic properties of the flavones, which are also used as supplement in the compost, have been studied. The purpose of this review is to evaluate the therapeutic effects of flavones in certain diseases. The positive effect of 16 flavones on health can be proven in many experimental studies and can be proven in the long run. Keywords: Functional nutrients, flavones, apigenin, luteolin, chrysin Introduction Polyphenolic flavonoids are among the affected by factors such as plant species, wide variety of phytochemicals found in the environment, genetics, light, maturity, human diet. Current studies reveal that harvest [3]. Flavones, a subset of dietary flavonoids are inversely related to flavonoids, form glycosylation and contain many cancers and age-related diseases [1]. -

Inhibitory Effect of Acacetin, Apigenin, Chrysin and Pinocembrin on Human Cytochrome P450 3A4
ORIGINAL SCIENTIFIC PAPER Croat. Chem. Acta 2020, 93(1), 33–39 Published online: August 03, 2020 DOI: 10.5562/cca3652 Inhibitory Effect of Acacetin, Apigenin, Chrysin and Pinocembrin on Human Cytochrome P450 3A4 Martin Kondža,1 Hrvoje Rimac,2,3 Željan Maleš,4 Petra Turčić,5 Ivan Ćavar,6 Mirza Bojić2,* 1 University of Mostar, Faculty of Pharmacy, Matice hrvatske bb, 88000 Mostar, Bosnia and Herzegovina 2 University of Zagreb, Faculty of Pharmacy and Biochemistry, Department of Medicinal Chemistry, A. Kovačića 1, 10000 Zagreb, Croatia 3 South Ural State University, Higher Medical and Biological School, Laboratory of Computational Modeling of Drugs, 454000 Chelyabinsk, Russian Federation 4 University of Zagreb, Faculty of Pharmacy and Biochemistry, Department of Pharmaceutical Botany, Schrottova 39, 10000 Zagreb, Croatia 5 University of Zagreb, Faculty of Pharmacy and Biochemistry, Department of Pharmacology, Domagojeva 2, 10000 Zagreb, Croatia 6 University of Mostar, Faculty of Medicine, Kralja Petra Krešimira IV bb, 88000 Mostar, Bosnia and Herzegovina * Corresponding author’s e-mail address: [email protected] RECEIVED: June 26, 2020 REVISED: July 28, 2020 ACCEPTED: July 30, 2020 Abstract: Cytochrome P450 3A4 is the most significant enzyme in metabolism of medications. Flavonoids are common secondary plant metabolites found in fruits and vegetables. Some flavonoids can interact with other drugs by inhibiting cytochrome P450 enzymes. Thus, the objective of this study was to determine inhibition kinetics of cytochrome P450 3A4 by flavonoids: acacetin, apigenin, chrysin and pinocembrin. For this purpose, testosterone was used as marker substrate, and generation of the 6β-hydroxy metabolite was monitored by high performance liquid chromatography coupled with diode array detector. -
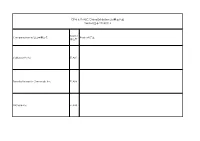
Cphi & P-MEC China Exhibition List展商名单version版本20180116
CPhI & P-MEC China Exhibition List展商名单 Version版本 20180116 Booth/ Company Name/公司中英文名 Product/产品 展位号 Carbosynth Ltd E1A01 Toronto Research Chemicals Inc E1A08 SiliCycle Inc. E1A10 SA TOURNAIRE E1A11 Indena SpA E1A17 Trifarma E1A21 LLC Velpharma E1A25 Anuh Pharma E1A31 Chemclone Industries E1A51 Hetero Labs Limited E1B09 Concord Biotech Limited E1B10 ScinoPharm Taiwan Ltd E1B11 Dongkook Pharmaceutical Co., Ltd. E1B19 Shenzhen Salubris Pharmaceuticals Co., Ltd E1B22 GfM mbH E1B25 Leawell International Ltd E1B28 DCS Pharma AG E1B31 Agno Pharma E1B32 Newchem Spa E1B35 APEX HEALTHCARE LIMITED E1B51 AMRI E1C21 Aarti Drugs Limited E1C25 Espee Group Innovators E1C31 Ruland Chemical Co., Ltd. E1C32 Merck Chemicals (Shanghai) Co., Ltd. E1C51 Mediking Pharmaceutical Group Ltd E1C57 珠海联邦制药股份有限公司/The United E1D01 Laboratories International Holdings Ltd. FMC Corporation E1D02 Kingchem (Liaoning) Chemical Co., Ltd E1D10 Doosan Corporation E1D22 Sunasia Co., Ltd. E1D25 Bolon Pharmachem Co., Ltd. E1D26 Savior Lifetec Corporation E1D27 Alchem International Pvt Ltd E1D31 Polish Investment and Trade Agency E1D57 Fischer Chemicals AG E1E01 NGL Fine Chem Limited E1E24 常州艾柯轧辊有限公司/ECCO Roller E1E25 Linnea SA E1E26 Everlight Chemical Industrial Corporation E1E27 HARMAN FINOCHEM E1E28 Zhechem Co Ltd E1F01 Midas Pharma GmbH Shanghai Representativ E1F03 Supriya Lifescience Ltd E1F10 KOA Shoji Co Ltd E1F22 NOF Corporation E1F24 上海贺利氏工业技术材料有限公司/Heraeus E1F26 Materials Technology Shanghai Ltd. Novacyl Asia Pacific Ltd E1F28 PharmSol Europe Limited E1F32 Bachem AG E1F35 Louston International Inc. E1F51 High Science Co Ltd E1F55 Chemsphere Technology Inc. E1F57a PharmaCore Biotech Co., Ltd. E1F57b Rockwood Lithium GmbH E1G51 Sarv Bio Labs Pvt Ltd E1G57 抗病毒类、抗肿瘤类、抗感染类和甾体类中间体、原料药和药物制剂及医药合约研发和加工服务 上海创诺医药集团有限公司/Shanghai Desano APIs and Finished products of ARV, Oncology, Anti-infection and Hormone drugs and E1H01 Pharmaceuticals Co., Ltd. -

Characterization of the CYP3A4 Enzyme Inhibition Potential of Selected Flavonoids
Article Characterization of the CYP3A4 Enzyme Inhibition Potential of Selected Flavonoids Martin Kondža 1 , Mirza Boji´c 2,* , Ivona Tomi´c 1, Željan Maleš 2 , Valentina Rezi´c 3 and Ivan Cavar´ 4 1 Faculty of Pharmacy, University of Mostar, Matice Hrvatske bb, 88000 Mostar, Bosnia and Herzegovina; [email protected] (M.K.); [email protected] (I.T.) 2 University of Zagreb Faculty of Pharmacy and Biochemistry, Ante Kovaˇci´ca1, 10000 Zagreb, Croatia; [email protected] 3 Farmavita d.o.o., Igmanska 5A, 71000 Sarajevo, Bosnia and Herzegovina; [email protected] 4 Faculty of Medicine, University of Mostar, Zrinskog Frankopana 34, 88000 Mostar, Bosnia and Herzegovina; [email protected] * Correspondence: [email protected]; Tel.: +385-1-4818-304 Abstract: Acacetin, apigenin, chrysin, and pinocembrin are flavonoid aglycones found in foods such as parsley, honey, celery, and chamomile tea. Flavonoids can act as substrates and inhibitors of the CYP3A4 enzyme, a heme containing enzyme responsible for the metabolism of one third of drugs on the market. The aim of this study was to investigate the inhibitory effect of selected flavonoids on the CYP3A4 enzyme, the kinetics of inhibition, the possible covalent binding of the inhibitor to the enzyme, and whether flavonoids can act as pseudo-irreversible inhibitors. For the determination of inhibition kinetics, nifedipine oxidation was used as a marker reaction. A hemochromopyridine test was used to assess the possible covalent binding to the heme, and incubation with dialysis was used in order to assess the reversibility of the inhibition. -

Natural Compounds to Counteract Testosterone Depletion in Aging
s & H oid orm er o t n S f a Rojas and Pandey, J Steroids Horm Sci 2014, 5:1 l o S l c a i Journal of DOI: 10.4172/2157-7536.1000e112 n e r n u c o e J ISSN: 2157-7536 Steroids & Hormonal Science Editorial Open Access Natural Compounds to Counteract Testosterone Depletion in Aging Daniela P Rojas and Akhilesh K Pandey* Garrison Institute on Aging, Department of Neurology, Texas Tech University Health Sciences Center, Lubbock, TX 79430, USA Aging is accompanied by many biological and physiological been seen that mutations in StAR protein result in sometimes lethal changes in the body, including the depletion of sex hormones such condition called, congenital lipoid adrenal hyperplasia, which involves as testosterone. Testosterone is an important sex hormone for many a blockage in the synthesis of steroid hormones, again expressing the physiological and biological functions in the body, and depletion of it importance of StAR protein function to produce testosterone [7]. StAR may cause changes that can be negative to the normal functioning of protein is expressed via a response to trophic hormone and it is cAMP- the body. Aging is associated with a gradual depletion of testosterone of mediated [8]. about 2-3% annually, starting from the age of 30 [1]. This depletion of With this information, one can conclude that aging is related the sex hormones has many implications in the physiological functions to a decrease in the levels of StAR protein and, thus, a decrease in in the body, such as effects in muscle mass, liver function and immune testosterone biosynthesis. -

Apigenin 520-36-5
SUMMARY OF DATA FOR CHEMICAL SELECTION Apigenin 520-36-5 BASIS OF NOMINATION TO THE CSWG Apigenin is brought to the attention of the CSWG because of a recent scientific article citing this flavonoid as a substance that can be metabolically activated to produce toxic prooxidant phenoxyl radicals. Pure apigenin is used primarily in research as a protein kinase inhibitor that may suppress tumor promotion and that has anti-proliferating effects on human breast cancer cells and inhibitory actions on MAP kinase. Apigenin is also one of several active ingredients in the popular herbal remedy, chamomile. Apigenin is found naturally in many fruits and vegetables, including apples and celery. It is found in several popular spices, including basil, oregano, tarragon, cilantro, and parsley. As a representative of flavonoids containing phenol B rings that may induce lipid peroxidation, apigenin is a candidate for testing. SELECTION STATUS ACTION BY CSWG: 12/12/00 Studies requested: Developmental toxicity Short-term tests for chromosomal aberrations Priority: None assigned Rationale/Remarks: Nomination based on concerns about apigenin’s potential to produce possibly toxic radicals and its estrogenic activity NCI will conduct a mouse lymphoma assay Apigenin 520-36-5 CHEMICAL IDENTIFICATION CAS Registry Number: 520-36-5 Chemical Abstracts Service Name: 4H-1-benzopyran-4-one,5,7-dihydroxy-2-(4- hydroxy-phenyl)- (9CI) Synonyms and Trade Names: Apigenin; apigenine; apigenol; chamomile; C.I. natural yellow 1; 2-(p-hydroxyphenyl)-5,7- dihydroxy-chromone; spigenin; 4',5,7- trihydroxyflavone Structural Class: Flavone Structure, Molecular Formula and Molecular Weight: OH HO O OH O C15H10O5 Mol. -

Progesterone Receptor Enhances Breast Cancer Cell Motility and Invasion Via Extranuclear Activation of Focal Adhesion Kinase
Endocrine-Related Cancer (2010) 17 431–443 Progesterone receptor enhances breast cancer cell motility and invasion via extranuclear activation of focal adhesion kinase Xiao-Dong Fu1,2*, Lorenzo Goglia1*, Angel Matias Sanchez1, Marina Flamini1, Maria S Giretti1, Veronica Tosi1, Andrea R Genazzani1 and Tommaso Simoncini1 1Molecular and Cellular Gynecological Endocrinology Laboratory (MCGEL), Department of Reproductive Medicine and Child Development, University of Pisa, Pisa 56100, Italy 2Department of Physiology, Zhongshan School of Medicine, Sun Yat-sen University, Guangzhou 510080, People’s Republic of China (Correspondence should be addressed to T Simoncini; Email: [email protected]) *(X-D Fu and L Goglia contributed equally to this work) Abstract While progesterone plays multiple roles in the process of breast development and differentiation, its role in breast cancer is less understood. We have shown previously that progestins stimulate breast cancer cell migration and invasion because of the activation of rapid signaling cascades leading to modifications in the actin cytoskeleton and cell membrane that are required for cell movement. In this study, we have investigated the effects of progesterone on the formation of focal adhesion (FA) complexes, which provide anchoring sites for cell attachment to the extracellular matrix during cell movement and invasion. In T47-D breast cancer cells, progesterone rapidly enhances FA kinase (FAK) phosphorylation at Tyr397 in a time- and concentration-dependent manner. As a result, exposure to progesterone leads to increased formation of FA complexes within specialized cell membrane protrusions. The cascade of events required for this phenomenon involves progesterone receptor interaction with the tyrosine kinase c-Src, which activates the phosphatidylinositol-3-kinase/Akt pathway and the small GTPase RhoA/Rho-associated kinase complex. -

Heat-Stabilised Rice Bran Consumption by Colorectal Cancer Survivors Modulates Stool Metabolite Profiles and Metabolic Networks: a Randomised Controlled Trial
Downloaded from British Journal of Nutrition (2017), 117, 1244–1256 doi:10.1017/S0007114517001106 © The Authors 2017. This is an Open Access article, distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike licence (http://creativecommons.org/licenses/by-nc-sa/4.0/), https://www.cambridge.org/core which permits non-commercial re-use, distribution, and reproduction in any medium, provided the same Creative Commons licence is included and the original work is properly cited. The written permission of Cambridge University Press must be obtained for commercial re-use. Heat-stabilised rice bran consumption by colorectal cancer survivors modulates stool metabolite profiles and metabolic networks: a randomised controlled trial . IP address: Dustin G. Brown1, Erica C. Borresen1, Regina J. Brown2,3 and Elizabeth P. Ryan1,3* 170.106.33.14 1Department of Environmental and Radiological Health Sciences, Colorado State University, Fort Collins, CO 80521, USA 2Division of Medical Oncology, University of Colorado School of Medicine, Aurora, CO 80045, USA , on 3University of Colorado Cancer Center, Aurora, CO 80045, USA 25 Sep 2021 at 23:07:56 (Submitted 28 October 2016 – Final revision received 28 March 2017 – Accepted 11 April 2017) Abstract Rice bran (RB) consumption has been shown to reduce colorectal cancer (CRC) growth in mice and modify the human stool microbiome. , subject to the Cambridge Core terms of use, available at Changes in host and microbial metabolism induced by RB consumption was hypothesised to modulate the stool metabolite profile in favour of promoting gut health and inhibiting CRC growth. The objective was to integrate gut microbial metabolite profiles and identify metabolic pathway networks for CRC chemoprevention using non-targeted metabolomics. -

Binding of Androgen- and Estrogen-Like Flavonoids to Their Cognate (Non)Nuclear Receptors: a Comparison by Computational Prediction
molecules Article Binding of Androgen- and Estrogen-Like Flavonoids to Their Cognate (Non)Nuclear Receptors: A Comparison by Computational Prediction Giulia D’Arrigo 1, Eleonora Gianquinto 1, Giulia Rossetti 2,3,4 , Gabriele Cruciani 5, Stefano Lorenzetti 6,* and Francesca Spyrakis 1,* 1 Department of Drug Science and Technology, University of Turin, Via Giuria 9, 10125 Turin, Italy; [email protected] (G.D.); [email protected] (E.G.) 2 Institute for Neuroscience and Medicine (INM-9) and Institute for Advanced Simulations (IAS-5) “Computational Biomedicine”, Forschungszentrum Jülich, 52425 Jülich, Germany 3 Jülich Supercomputing Center (JSC), Forschungszentrum Jülich, 52425 Jülich, Germany 4 Department of Neurology, RWTH, Aachen University, 52074 Aachen, Germany; [email protected] 5 Department of Chemistry, Biology and Biotechnology, University of Perugia, 06123 Perugia, Italy; [email protected] 6 Istituto Superiore di Sanità (ISS), Department of Food Safety, Nutrition and Veterinary Public Health, Viale Regina Elena 299, 00161 Rome, Italy * Correspondence: [email protected] (S.L.); [email protected] (F.S.) Abstract: Flavonoids are plant bioactives that are recognized as hormone-like polyphenols because of Citation: D’Arrigo, G.; Gianquinto, their similarity to the endogenous sex steroids 17β-estradiol and testosterone, and to their estrogen- E.; Rossetti, G.; Cruciani, G.; and androgen-like activity. Most efforts to verify flavonoid binding to nuclear receptors (NRs) and Lorenzetti, S.; Spyrakis, F. Binding of explain their action have been focused on ERα, while less attention has been paid to other nuclear Androgen- and Estrogen-Like and non-nuclear membrane androgen and estrogen receptors. Here, we investigate six flavonoids Flavonoids to Their Cognate (apigenin, genistein, luteolin, naringenin, quercetin, and resveratrol) that are widely present in fruits (Non)Nuclear Receptors: A and vegetables, and often used as replacement therapy in menopause.