Organophosphate Ester Induced Chronic Neurotoxicity (Opicn)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Hyperactivity and Seizure Induced by Tricresyl Phosphate Are Isomer
TOXICOLOGICAL SCIENCES, 2021, 1–15 doi: 10.1093/toxsci/kfab006 Research Article Downloaded from https://academic.oup.com/toxsci/advance-article/doi/10.1093/toxsci/kfab006/6112059 by guest on 12 February 2021 Hyperactivity and Seizure Induced by Tricresyl Phosphate Are Isomer Specific and Not Linked to Phenyl Valerate-Neuropathy Target Esterase Activity Inhibition in Zebrafish Anja Knoll-Gellida, Leslie E. Dubrana, Laure M. Bourcier, Theo Merce, Gaelle€ Gruel, Magalie Soares, and Patrick J. Babin 1 Department of Life and Health Sciences, INSERM, Maladies Rares: Gen etique et Metabolisme (MRGM), U1211, Universite de Bordeaux, F-33615 Pessac, France Anja Knoll-Gellida and Leslie E. Dubrana contributed equally to this study. 1To whom correspondence should be addressed at: Laboratoire Maladies Rares: Gen etique et Metabolisme (MRGM), Universite de Bordeaux, Allee Geoffroy St-Hilaire, Bat. B2, 2e`meetage, CS 50023, 33615 Pessac Cedex, France. E-mail: [email protected] ABSTRACT Environmental exposure to tricresyl phosphate (TCP) may lead to severe neurotoxic effects, including organophosphate (OP)-induced delayed neuropathy. TCP has three symmetric isomers, distinguished by the methyl group position on the aromatic ring system. One of these isomers, tri-ortho-cresyl phosphate (ToCP), has been reported for years as a neuropathic OP, targeting neuropathic target esterase (NTE/PNPLA6), but its mode of toxic action had not been fully elucidated. Zebrafish eleuthero-embryo and larva were used to characterize the differential action of the TCP isomers. The symmetric isomers inhibited phenyl valerate (PV)-NTE enzymatic activity in vivo with different IC50, while no effect was observed on acetylcholinesterase activity. Moreover, the locomotor behavior was also affected by tri-para-cresyl phosphate and tri- meta-cresyl phosphate, only ToCP exposure led to locomotor hyperactivity lasting several hours, associated with defects in the postural control system and an impaired phototactic response, as revealed by the visual motor response test. -

Alphabetical Index of Substances and Articles
ALPHABETICAL INDEX OF SUBSTANCES AND ARTICLES - 355 - NOTES TO THE INDEX 1. This index is an alphabetical list of the substances and articles which are listed in numerical order in the Dangerous Goods List in Chapter 3.2. 2. For the purpose of determining the alphabetical order the following information has been ignored even when it forms part of the proper shipping name: numbers; Greek letters; the abbreviations “sec” and “tert”; and the letters “N” (nitrogen), “n” (normal), “o” (ortho) “m” (meta), “p” (para) and “N.O.S.” (not otherwise specified). 3. The name of a substance or article in block capital letters indicates a proper shipping name. 4. The name of a substance or article in block capital letters followed by the word “see” indicates an alternative proper shipping name or part of a proper shipping name (except for PCBs). 5. An entry in lower case letters followed by the word “see” indicates that the entry is not a proper shipping name; it is a synonym. 6. Where an entry is partly in block capital letters and partly in lower case letters, the latter part is considered not to be part of the proper shipping name. 7. A proper shipping name may be used in the singular or plural, as appropriate, for the purposes of documentation and package marking. - 356 - INDEX Name and description Class UN No. Name and description Class UN No. Accumulators, electric, see 4.3 3292 Acid mixture, nitrating acid, see 8 1796 8 2794 8 2795 Acid mixture, spent, nitrating acid, see 8 1826 8 2800 8 3028 Acraldehyde, inhibited, see 6.1 1092 ACETAL 3 1088 -

Structural Dynamics of Acetylcholinesterase and Its Implications in Reactivators Design Gianluca Santoni
Structural dynamics of acetylcholinesterase and its implications in reactivators design Gianluca Santoni To cite this version: Gianluca Santoni. Structural dynamics of acetylcholinesterase and its implications in reactivators design. Biomolecules [q-bio.BM]. Université Grenoble Alpes, 2015. English. NNT : 2015GREAY019. tel-01212481 HAL Id: tel-01212481 https://tel.archives-ouvertes.fr/tel-01212481 Submitted on 6 Oct 2015 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. THÈSE Pour obtenir le grade de DOCTEUR DE L’UNIVERSITÉ DE GRENOBLE Spécialité : Physique pour les sciences du vivant Arrêté ministériel : 7 Aout 2006 Présentée par Gianluca SANTONI Thèse dirigée par Martin WEIK et codirigée par Florian NACHON préparée au sein de l’Institut de Biologie Structurale de Grenoble et de l’école doctorale de physique Structural dynamics of acetyl- cholinesterase and its implications in reactivator design Thèse soutenue publiquement le 30/01/2015, devant le jury composé de : Dr. Yves Bourne Directeur de recherche CNRS, AFMB Marseille, Rapporteur Dr. Etienne Derat Maitre de conference, Université Pierre et Marie Curie, Paris, Rapporteur Prof. Pierre-Yves Renard Professeur, Université de Normandie, Rouen, Examinateur Prof. Israel Silman Professeur, Weizmann Institute of Science,Rehovot, Examinateur Dr. -
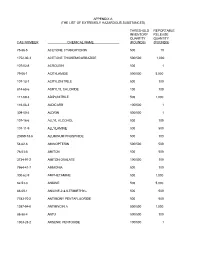
The List of Extremely Hazardous Substances)
APPENDIX A (THE LIST OF EXTREMELY HAZARDOUS SUBSTANCES) THRESHOLD REPORTABLE INVENTORY RELEASE QUANTITY QUANTITY CAS NUMBER CHEMICAL NAME (POUNDS) (POUNDS) 75-86-5 ACETONE CYANOHYDRIN 500 10 1752-30-3 ACETONE THIOSEMICARBAZIDE 500/500 1,000 107-02-8 ACROLEIN 500 1 79-06-1 ACRYLAMIDE 500/500 5,000 107-13-1 ACRYLONITRILE 500 100 814-68-6 ACRYLYL CHLORIDE 100 100 111-69-3 ADIPONITRILE 500 1,000 116-06-3 ALDICARB 100/500 1 309-00-2 ALDRIN 500/500 1 107-18-6 ALLYL ALCOHOL 500 100 107-11-9 ALLYLAMINE 500 500 20859-73-8 ALUMINUM PHOSPHIDE 500 100 54-62-6 AMINOPTERIN 500/500 500 78-53-5 AMITON 500 500 3734-97-2 AMITON OXALATE 100/500 100 7664-41-7 AMMONIA 500 100 300-62-9 AMPHETAMINE 500 1,000 62-53-3 ANILINE 500 5,000 88-05-1 ANILINE,2,4,6-TRIMETHYL- 500 500 7783-70-2 ANTIMONY PENTAFLUORIDE 500 500 1397-94-0 ANTIMYCIN A 500/500 1,000 86-88-4 ANTU 500/500 100 1303-28-2 ARSENIC PENTOXIDE 100/500 1 THRESHOLD REPORTABLE INVENTORY RELEASE QUANTITY QUANTITY CAS NUMBER CHEMICAL NAME (POUNDS) (POUNDS) 1327-53-3 ARSENOUS OXIDE 100/500 1 7784-34-1 ARSENOUS TRICHLORIDE 500 1 7784-42-1 ARSINE 100 100 2642-71-9 AZINPHOS-ETHYL 100/500 100 86-50-0 AZINPHOS-METHYL 10/500 1 98-87-3 BENZAL CHLORIDE 500 5,000 98-16-8 BENZENAMINE, 3-(TRIFLUOROMETHYL)- 500 500 100-14-1 BENZENE, 1-(CHLOROMETHYL)-4-NITRO- 500/500 500 98-05-5 BENZENEARSONIC ACID 10/500 10 3615-21-2 BENZIMIDAZOLE, 4,5-DICHLORO-2-(TRI- 500/500 500 FLUOROMETHYL)- 98-07-7 BENZOTRICHLORIDE 100 10 100-44-7 BENZYL CHLORIDE 500 100 140-29-4 BENZYL CYANIDE 500 500 15271-41-7 BICYCLO[2.2.1]HEPTANE-2-CARBONITRILE,5- -

Chemical Compatibility of the Tubing Materials
Chemical Compatibility of the Tubing Materials 1: excellent; 2: good; 3: fair; 4: not recommended Tygon Tygon ST PharMed Tygon HC Tygon Tygon SI Silicone Norprene Flurane LFL R-3603 F-4040-A MH Silicone Peroxide A-60-G F-5500-A Name 2075 Platinum (Viton) Acetaldehyde 4 4 4 4 3 3 3 4 4 Acetamide, 67% in w 4 4 2 4 1 1 1 2 4 Acetate Solvents 4 4 2 4 4 4 4 2 4 Acetic Acid, 10% in w 1 1 1 1 1 1 1 1 4 Acetic Acid, 50-60% in w 1 1 2 1 1 1 1 2 4 Acetic Acid, Glacial, 100% 4 4 2 4 1 4 4 2 4 Acetic Anhydride 4 4 1 4 1 1 1 1 4 Acetone 4 4 4 4 2 3 3 4 4 Acetonitrile 4 4 4 4 4 4 4 4 2 Acetyl Bromide 4 4 3 4 4 4 4 3 4 Acetyl Chloride 4 4 3 4 4 4 4 3 4 Acetylene Gas 1 1 1 1 1 1 1 1 1 Acrylonitrile 4 4 4 4 4 4 4 4 2 Adipic Acid, 100% in alc 4 4 2 3 4 4 4 2 4 Air 1 1 1 1 1 1 1 1 1 Alcohols General 4 4 1 2 1 2 2 1 4 Aliphatic Hydrocarbons 4 4 4 2 4 4 4 4 2 Allyl Alcohol 4 4 3 1 1 4 4 3 1 Alum, 5% in w 1 1 1 1 1 1 1 1 1 Aluminum Chloride, 53% in w 1 1 1 1 1 1 1 1 1 Aluminum Hydroxide, 2% in w 1 1 1 1 1 1 1 1 1 Aluminum Salts 1 1 1 1 1 1 1 1 1 Aluminum Sulfate, 50% in w 1 1 1 1 1 1 1 1 1 Amines 443444434 Ammonia Gas 1 1 1 1 1 4 4 1 4 Ammonia, Anhydrous Liquid 2 2 1 2 2 4 4 2 4 Ammonium Acetate, 45% in w 1 1 1 1 1 1 1 1 4 Ammonium Carbonate, 20% in w 1 1 1 1 1 1 1 1 1 Ammonium Hydroxide, 30% in w 1 1 1 3 1 4 4 1 4 Ammonium Hydroxide, 5-10% in w 1 1 1 2 1 4 4 1 4 Ammonium Persulfate, 30% in w 1 1 1 1 1 1 1 1 1 Ammonium Salts 1 1 1 1 1 1 1 1 1 Ammonium Sulfate, 30% in w 1 1 1 1 1 1 1 1 1 Amyl Acetate 4 4 2 4 4 4 4 2 4 Amyl Alcohol 4 4 4 1 1 4 4 4 1 Amyl Chloride -

744 Hydrolysis of Chiral Organophosphorus Compounds By
[Frontiers in Bioscience, Landmark, 26, 744-770, Jan 1, 2021] Hydrolysis of chiral organophosphorus compounds by phosphotriesterases and mammalian paraoxonase-1 Antonio Monroy-Noyola1, Damianys Almenares-Lopez2, Eugenio Vilanova Gisbert3 1Laboratorio de Neuroproteccion, Facultad de Farmacia, Universidad Autonoma del Estado de Morelos, Morelos, Mexico, 2Division de Ciencias Basicas e Ingenierias, Universidad Popular de la Chontalpa, H. Cardenas, Tabasco, Mexico, 3Instituto de Bioingenieria, Universidad Miguel Hernandez, Elche, Alicante, Spain TABLE OF CONTENTS 1. Abstract 2. Introduction 2.1. Organophosphorus compounds (OPs) and their toxicity 2.2. Metabolism and treatment of OP intoxication 2.3. Chiral OPs 3. Stereoselective hydrolysis 3.1. Stereoselective hydrolysis determines the toxicity of chiral compounds 3.2. Hydrolysis of nerve agents by PTEs 3.2.1. Hydrolysis of V-type agents 3.3. PON1, a protein restricted in its ability to hydrolyze chiral OPs 3.4. Toxicity and stereoselective hydrolysis of OPs in animal tissues 3.4.1. The calcium-dependent stereoselective activity of OPs associated with PON1 3.4.2. Stereoselective hydrolysis commercial OPs pesticides by alloforms of PON1 Q192R 3.4.3. PON1, an enzyme that stereoselectively hydrolyzes OP nerve agents 3.4.4. PON1 recombinants and stereoselective hydrolysis of OP nerve agents 3.5. The activity of PTEs in birds 4. Conclusions 5. Acknowledgments 6. References 1. ABSTRACT Some organophosphorus compounds interaction of the racemic OPs with these B- (OPs), which are used in the manufacturing of esterases (AChE and NTE) and such interactions insecticides and nerve agents, are racemic mixtures have been studied in vivo, ex vivo and in vitro, using with at least one chiral center with a phosphorus stereoselective hydrolysis by A-esterases or atom. -

Environmental Health Criteria 110 Tricresyl Phosphate
Environmental Health Criteria 110 Tricresyl phosphate Please note that the layout and pagination of this web version are not identical with the printed version. Tricresyl phosphate (EHC 110, 1990) INTERNATIONAL PROGRAMME ON CHEMICAL SAFETY ENVIRONMENTAL HEALTH CRITERIA 110 TRICRESYL PHOSPHATE This report contains the collective views of an international group of experts and does not necessarily represent the decisions or the stated policy of the United Nations Environment Programme, the International Labour Organisation, or the World Health Organization. Published under the joint sponsorship of the United Nations Environment Programme, the International Labour Organisation, and the World Health Organization World Health Orgnization Geneva, 1990 The International Programme on Chemical Safety (IPCS) is a joint venture of the United Nations Environment Programme, the International Labour Organisation, and the World Health Organization. The main objective of the IPCS is to carry out and disseminate evaluations of the effects of chemicals on human health and the quality of the environment. Supporting activities include the development of epidemiological, experimental laboratory, and risk-assessment methods that could produce internationally comparable results, and the development of manpower in the field of toxicology. Other activities carried out by the IPCS include the development of know-how for coping with chemical accidents, coordination of laboratory testing and epidemiological studies, and promotion of research on the mechanisms of the biological action of chemicals. WHO Library Cataloguing in Publication Data Tricresyl phosphate. (Environmental health criteria ; 110) 1.Tritolyl phosphates - adverse effects 2.Tritolyl phosphates - toxicity I.Series Page 1 of 84 Tricresyl phosphate (EHC 110, 1990) ISBN 92 4 157110 1 (NLM Classification: QV 627) ISSN 0250-863X The World Health Organization welcomes requests for permission to reproduce or translate its publications, in part or in full. -

NIOSH Method 5600: Organophosphorus Pesticides
ORGANOPHOSPHORUS PESTICIDES 5600 Formula: Table 1 MW: Table 1 CAS: Table 1 RTECS: Table 1 METHOD: 5600, Issue 1 EVALUATION: FULL Issue 1: 15 August 1994 OSHA : Table 2 PROPERTIES: Table 3 NIOSH: Table 2 ACGIH: Table 2 SYNONYMS: Table 4 SAMPLING MEASUREMENT SAMPLER: FILTER/SOLID SORBENT TUBE (OVS-2 tube: TECHNIQUE: GC, FLAME PHOTOMETRIC DETECTION 13-mm quartz filter; XAD-2, 270 mg/140 mg) (FPD) FLOW RATE: 0.2 to 1 L/min ANALYTE: organophosphorus pesticides, Table 1 VOL-MIN: 12 L EXTRACTION: 2-mL 90% toluene/10% acetone solution -MAX: 240 L; 60 L (Malathion, Ronnel) INJECTION SHIPMENT: cap both ends of tube VOLUME: 1-2 µL SAMPLE TEMPERATURE STABILITY: at least 10 days at 25 °C -INJECTION: 240 °C at least 30 days at 0 °C -DETECTOR: 180 °C to 215 °C (follow manufacturer's recommendation) BLANKS: 2 to 10 field blanks per set -COLUMN: Table 6 CARRIER GAS: He at 15 psi (104 kPa) ACCURACY COLUMN: fused silica capillary column; Table 6 RANGE STUDIED: Table 5, Column A DETECTOR: FPD (phosphorus mode) ACCURACY: Table 5, Column B CALIBRATION: standard solutions of organophosphorus compounds in toluene BIAS: Table 5, Column C RANGE: Table 8, Column C ˆ OVERALL PRECISION (S rT): Table 5, Column D ESTIMATED LOD: Table 8, Column F PRECISION (S r): Table 5, Column E APPLICABILITY: The working ranges are listed in Table 5. They cover a range of 1/10 to 2 times the OSHA PELs. This INTERFERENCES: Several organophosphates may co-elute method also is applicable to STEL measurements using 12-L with either target analyte or internal standard causing samples. -

Quantitation of Ortho-Cresyl Phosphate Adducts to Butyrylcholinesterase in Human Serum by Immunomagnetic- UHPLC-MS/MS
HHS Public Access Author manuscript Author ManuscriptAuthor Manuscript Author J Mass Spectrom Manuscript Author . Author Manuscript Author manuscript; available in PMC 2017 December 04. Published in final edited form as: J Mass Spectrom. 2015 April ; 50(4): 683–692. doi:10.1002/jms.3576. Quantitation of ortho-Cresyl Phosphate Adducts to Butyrylcholinesterase in Human Serum by Immunomagnetic- UHPLC-MS/MS Darryl Johnson1, Melissa D. Carter2,*, Brian S. Crow2, Samantha L. Isenberg1, Leigh Ann Graham1, H. Akin Erol1, Caroline M. Watson1, Brooke G. Pantazides2, Marcel J. van der Schans3, Jan P. Langenberg3, Daan Noort3, Thomas A. Blake2, Jerry D. Thomas2, and Rudolph C. Johnson2 1Oak Ridge Institute for Science and Education, Centers for Disease Control and Prevention, Atlanta, GA, USA 2Division of Laboratory Sciences, National Center for Environmental Health, Centers for Disease Control and Prevention, Atlanta, GA, USA 3The Netherlands Organization for Applied Scientific Research (TNO), Technical Sciences, CBRN Protection, Rijswijk, The Netherlands Abstract Tri-ortho-cresyl phosphate (ToCP) is an anti-wear, flame retardant additive used in industrial lubricants, hydraulic fluids, and gasoline. The neurotoxic effects of ToCP arise from the liver- activated metabolite 2-(o-cresyl)-4H-1,3,2-benzodioxaphosphoran-2-one (cresyl saligenin phosphate or CBDP), which inhibits esterase enzymes including butyrylcholinesterase (BChE). Following BChE adduction, CBDP undergoes hydrolysis to form the aged adduct ortho-cresyl phosphoserine (oCP-BChE), thus providing a biomarker of CBDP exposure. Previous studies have identified ToCP in aircraft cabin and cockpit air, but assessing human exposure has been hampered by the lack of a laboratory assay to confirm exposure. This work presents the development of an immunomagnetic-UHPLC-MS/MS method for the quantitation of unadducted BChE and the long- term CBDP biomarker, oCP-BChE, in human serum. -

2019 Minnesota Chemicals of High Concern List
Minnesota Department of Health, Chemicals of High Concern List, 2019 Persistent, Bioaccumulative, Toxic (PBT) or very Persistent, very High Production CAS Bioaccumulative Use Example(s) and/or Volume (HPV) Number Chemical Name Health Endpoint(s) (vPvB) Source(s) Chemical Class Chemical1 Maine (CA Prop 65; IARC; IRIS; NTP Wood and textiles finishes, Cancer, Respiratory 11th ROC); WA Appen1; WA CHCC; disinfection, tissue 50-00-0 Formaldehyde x system, Eye irritant Minnesota HRV; Minnesota RAA preservative Gastrointestinal Minnesota HRL Contaminant 50-00-0 Formaldehyde (in water) system EU Category 1 Endocrine disruptor pesticide 50-29-3 DDT, technical, p,p'DDT Endocrine system Maine (CA Prop 65; IARC; IRIS; NTP PAH (chem-class) 11th ROC; OSPAR Chemicals of Concern; EuC Endocrine Disruptor Cancer, Endocrine Priority List; EPA Final PBT Rule for 50-32-8 Benzo(a)pyrene x x system TRI; EPA Priority PBT); Oregon P3 List; WA Appen1; Minnesota HRV WA Appen1; Minnesota HRL Dyes and diaminophenol mfg, wood preservation, 51-28-5 2,4-Dinitrophenol Eyes pesticide, pharmaceutical Maine (CA Prop 65; IARC; NTP 11th Preparation of amino resins, 51-79-6 Urethane (Ethyl carbamate) Cancer, Development ROC); WA Appen1 solubilizer, chemical intermediate Maine (CA Prop 65; IARC; IRIS; NTP Research; PAH (chem-class) 11th ROC; EPA Final PBT Rule for 53-70-3 Dibenzo(a,h)anthracene Cancer x TRI; WA PBT List; OSPAR Chemicals of Concern); WA Appen1; Oregon P3 List Maine (CA Prop 65; NTP 11th ROC); Research 53-96-3 2-Acetylaminofluorene Cancer WA Appen1 Maine (CA Prop 65; IARC; IRIS; NTP Lubricant, antioxidant, 55-18-5 N-Nitrosodiethylamine Cancer 11th ROC); WA Appen1 plastics stabilizer Maine (CA Prop 65; IRIS; NTP 11th Pesticide (EPA reg. -

Environmental and Disposal Liabilities Identification, Documentation and Reporting Handbook V 3.0
Environmental and Disposal Liabilities Identification, Documentation And Reporting Handbook v 3.0 DECEMBER 2011 Environmental and Disposal Liabilities Identification, Documentation and Reporting Handbook v3.0 TABLE OF CONTENTS Page 1.0 INTRODUCTION .............................................................................................................. 1 1.1 Applicable Standards and Guidance ............................................................................... 1 1.2 Definitions ...................................................................................................................... 3 1.3 Responsibilities ............................................................................................................... 6 2.0 EDL IDENTIFICATION .................................................................................................... 7 2.1 Environmental Location of Concern............................................................................... 8 2.2 Deconstruction and Renovation Activities ..................................................................... 9 2.3 Environmental and Disposal Liabilities ........................................................................ 10 3.0 LIABILITY STATUS ...................................................................................................... 12 3.1 Probable ........................................................................................................................ 12 3.2 Reasonably Possible .................................................................................................... -

Genetic Variability and Sensitivity to Organophosphate Exposures
Appendix RAC-GWVI Meeting Minutes Presentation 5 - Furlong November 2-3, 2009 Page 109 of 269 Genetic Variability and Sensitivity to Organophosphate Exposures Research Advisory Committee on Gulf War Veterans Illnesses Meeting November 2, 2009 Clement E. Furlong, Research Professor Departments of Medicine - Division of Medical Genetics & Genome Sciences Goals of This Presentation The purpose of this brief presentation is to share with you what we have learned about organophosphate (OP) exposures and the consequences of genetic variability in modulating these exposures. One topic will be the role of plasma paraoxonase (PON1) in protecting against exposure to organophosphorus insecticides, particularly diazinon/diazoxon and chlorpyrifos/chlorpyrifos oxon and the consequences of genetic variability in modulating mixed OP exposures PON1 is a high density lipoprotein (HDL) associated enzyme of 354 amino acids that plays a significant role in the detoxication of the highly toxic OP metabolites diazoxon and chlorpyrifos oxon. The role of animal models in understanding the consequences of gene/environment interactions will also be discussed. Research on biomarkers of exposure, sensitivity and disease will also be discussed. Appendix RAC-GWVI Meeting Minutes Presentation 5 - Furlong November 2-3, 2009 Page 110 of 269 Biomarkers • Biomarkers of susceptibility Why are some individuals more susceptible than others to a given exposure? • Biomarkers of exposure How do you know if you have been exposed to a given toxicant (e.g., OP insecticide or tricresyl