Fossil Cenozoic Crassatelline Bivalves from Peru: New Species and Generic Insights
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Scottsdale Arabian Horse Show Feb 16 2017 Result Back# Horse Name
Results by Class: Scottsdale Arabian Horse Show Feb 16 2017 Result Back# Horse Name / Sire x Dam Owner / Breeder Handler Page: 1 Class#: 1G Arabian Hunter Pleasure JTR 17-18yrs 16 Shown Judge:Terri Delbridge Jody LaSalle Lisa Jo White 1 1071 C HONDO Wolf Springs Ranches Inc Anna Redmond ENZO X ENCHANTES BEY Bill Cunningham 2 1685 JUSTTINIAN Lauren Ambrosini Lauren Ambrosini JUSTIFY X SHAHMAICA BEY Robert C Or Janene M Boggs 3 1871 REDEMPTION H Alyssa or Marlene Tomusiak Alyssa Tomusiak MILLENNIUM LOA X JORDONA Peggy Heathcott 4 1030 SF AHEIR OVERDRESSED Michelle Kurth or Tiffany Kurth Madison Schwanz AFIRES HEIR X SF ALEXIS Springwater Farms Arabians LLC 5 284 PALADIN LL LMJ Investments LLC Julia Nastri MAGNUM CHALL HVP X NV GYPSY Leslie A Lurken DANCER 6 1972 CP GUNCYN ROSES Kirsten Luedde Kirsten Luedde CYTOSK X BARBS MIST Cal Poly Pomona 7 1750 KAYCEE OF JEWYLLS LAS Kathryn M Schwartz Abigail Buschette SHOWKAYCE X KHNIGHT JEWYLL LAS Lavoy Shepherd 8 1442 JUSTA HOOT Bailey and Jessica Mirmelli Bailey Mirmelli JUSTAFIRE DGL X FSR BARBARA Michael J or Victoria J Mangan 9 1759 ATLANTIS SC Charles or Christine or Adam Rickart Mikenna Laventure ODYSSEY SC X CARISMRETA Sparkle Creek LLC 10 753 KYROS ROMEO Wendy or Kaitlyn Ruonavaara Kaitlyn Ruonavaara DA VALENTINO X SR MAGIC MERCY Charlene L Strong Class#: 1H Arabian Hunter Pleasure JTR 15-16yrs 16 Shown Judge:Terri Delbridge Jody LaSalle Lisa Jo White 1 1043 DA SOVEREIGN Emma or Jody Freeland Emma Freeland SUNDANCE KID V X DA FAITH Dolorosa Arabians LTD 2 1881 ARAMIS LBA+ Ali Brady and Matthew -

Jacksonville, Florida 1998 Odmds Benthic Community Assessment
JACKSONVILLE, FLORIDA 1998 ODMDS BENTHIC COMMUNITY ASSESSMENT Submitted to U.S. Environmental Protection Agency, Region 4 61 Forsyth St. Atlanta, Georgia 30303 Prepared by Barry A. Vittor & Associates, Inc. 8060 Cottage Hill Rd. Mobile, Alabama 36695 (334) 633-6100 November 1999 TABLE OF CONTENTS LIST OF TABLES ………………………………………….……………………………3 LIST OF FIGURES ……………………..………………………………………………..4 1.0 INTRODUCTION ………..…………………………………………………………..5 2.0 METHODS ………..…………………………………………………………………..5 2.1 Sample Collection And Handling ………………………………………………5 2.2 Macroinfaunal Sample Analysis ……………………………………………….6 3.0 DATA ANALYSIS METHODS ……..………………………………………………6 3.1 Assemblage Analyses ..…………………………………………………………6 3.2 Faunal Similarities ……………………………………………………….…….8 4.0 HABITAT CHARACTERISTICS ……………………………………………….…8 5.0 BENTHIC COMMUNITY CHARACTERIZATION ……………………………..9 5.1 Faunal Composition, Abundance, And Community Structure …………………9 5.2 Numerical Classification Analysis …………………………………………….10 5.3 Taxa Assemblages …………………………………………………………….11 6.0 1995 vs 1998 COMPARISONS ……………………………………………………..11 7.0 SUMMARY ………………………………………………………………………….13 8.0 LITERATURE CITED ……………………………………………………………..16 2 LIST OF TABLES Table 1. Station locations for the Jacksonville, Florida ODMDS, June 1998. Table 2. Sediment data for the Jacksonville, Florida ODMDS, June 1998. Table 3. Summary of abundance of major taxonomic groups for the Jacksonville, Florida ODMDS, June 1998. Table 4. Abundance and distribution of major taxonomic groups at each station for the Jacksonville, Florida ODMDS, June 1998. Table 5. Abundance and distribution of taxa for the Jacksonville, Florida ODMDS, June 1998. Table 6. Percent abundance of dominant taxa (> 5% of the total assemblage) for the Jacksonville, Florida ODMDS, June 1998. Table 7. Summary of assemblage parameters for the Jacksonville, Florida ODMDS stations, June 1998. Table 8. Analysis of variance table for density differences between stations for the Jacksonville, Florida ODMDS stations, June 1998. -
Field Identification Guide to the Living Marine Resources In
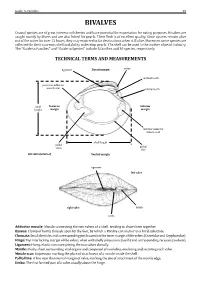
Guide to Families 29 BIVALVES Coastal species are of great interest to fisheries and have potential for exportation for eating purposes. Bivalves are caught mainly by divers and are also fished for pearls. Their flesh is of excellent quality. Since oysters remain alive out of the water for over 12 hours, they may exported to far destinations when still alive. Moreover, some species are collected for their nacreous shell and ability to develop pearls. The shell can be used in the mother of pearl industry. The “Guide to Families’’ andTECHNICAL ‘‘Guide to Species’’ TERMS include 5AND families MEASUREMENTS and 10 species, respectively. ligament Dorsal margin umbo posterior adductor cardinal tooth muscle scar lateral tooth Posterior Anterior margin margin shell height anterior adductor muscle scar pallial sinus pallial shell length line left valve (interior) Ventral margin ligament left valve right valve lunule umbo Adductor muscle: Byssus: Chomata: Muscle connecting the two valves of a shell, tending to draw them together. Hinge: Clump of horny threads spun by the foot, by which a Bivalve can anchor to a hard substrate. Ligament: Small denticles and corresponding pits located on the inner margin of the valves (Ostreidae and Gryphaeidae). Mantle: Top interlocking margin of the valves, often with shelly projections (teeth) and corresponding recesses (sockets). Muscle scar: Horny, elastic structure joining the two valves dorsally. Pallial line: Fleshy sheet surrounding vital organs and composed of two lobes, one lining and secreting each valve. Umbo: Impression marking the place of attachment of a muscle inside the shell. A line near the internal margin of valve, marking the site of attachment of the mantle edge. -

Freshwater Mussels of the Pacific Northwest
Freshwater Mussels of the Pacifi c Northwest Ethan Nedeau, Allan K. Smith, and Jen Stone Freshwater Mussels of the Pacifi c Northwest CONTENTS Part One: Introduction to Mussels..................1 What Are Freshwater Mussels?...................2 Life History..............................................3 Habitat..................................................5 Role in Ecosystems....................................6 Diversity and Distribution............................9 Conservation and Management................11 Searching for Mussels.............................13 Part Two: Field Guide................................15 Key Terms.............................................16 Identifi cation Key....................................17 Floaters: Genus Anodonta.......................19 California Floater...................................24 Winged Floater.....................................26 Oregon Floater......................................28 Western Floater.....................................30 Yukon Floater........................................32 Western Pearlshell.................................34 Western Ridged Mussel..........................38 Introduced Bivalves................................41 Selected Readings.................................43 www.watertenders.org AUTHORS Ethan Nedeau, biodrawversity, www.biodrawversity.com Allan K. Smith, Pacifi c Northwest Native Freshwater Mussel Workgroup Jen Stone, U.S. Fish and Wildlife Service, Columbia River Fisheries Program Offi ce, Vancouver, WA ACKNOWLEDGEMENTS Illustrations, -

Fossil Crustacea Atlantic and Gulf Coastal Plain
LIBRARY VJUSHMAN LABORATORY GE0WG1UAL UOVlkilk Ot AMERICA SPECIAL PAPERS NUMBER 2 FOSSIL CRUSTACEA OF THE ATLANTIC AND GULF COASTAL PLAIN BY MARY J. RATHBUN INVERTEBRATE \ ZOOLOGY % jDrustacea C | B R ft R-Y Wmi V CftUSTAOE* V PUBLISHED BY THE SOCIETY 14)35 COUNCIL, 1935 President: NEVIN M. FENNEMAN, Cincinnati, Ohio Past President: W. EL COLLINS, Ottawa, Canada Vice-Presidents: EDSON S. BASTIN, Chicago, 111. JOHN B. REESIDE, JR., Washington, D. C. DONNEL F. HEWETT, Washington, D. C. AUSTIN F. ROGERS, Stanford University, Cal. Secretary: CHARLES P. BERKEY, 419 W. 117th Street, New York, N. Y. Treasurer: EDWARD B. MATHEWS, Johns Hopkins University, Baltimore, Md. Councilors: (Term expires 1935) FRANK F. GROUT, Minneapolis, Minn. W. 0. HOTCHKISS, Houghton, Mich. JOSEPH STANLEY-BROWN, Kew Gardens, N. Y. (Term expires 1936) F. W. DEWOLF, Urbana, 111. D. H. MCLAUGHLIN, Cambridge, Mass. ADOLPH KNOPF, New Haven, Conn. (Term expires 1937) WALTER H. BUCHER, Cincinnati, Ohio RUSSELL S. KNAPPEN, Tulsa, Okla. E. L. BRUCE, Kingston, Ont., Canada GEOLOGICAL SOCIETY OF AMERICA SPECIAL PAPERS NUMBER 2 FOSSIL CRUSTACEA OF THE ATLANTIC AND GULF COASTAL PLAIN BY MARY J. RATHBUN L I B R a R Y pep^ Of O^UcTtkUEA ilDBRAT L ZOOLOGY Crustacea PUBLISHED BY THE SOCIETY 1935 WAVERLY PRESS, INC. BALTIMORE, MD. The Special Papers of The Geological Society of America are made possible through the bequest of Richard Alexander Fullerton Penrose, Jr. CONTENTS INTRODUCTION Statement of the Problem Sources of Material Unusual Extensions of Range Mingling of Cretaceous with Eocene Correlation with European Forms New Additions LIST OF SPECIES AND DISTRIBUTION DETAILED DESCRIPTION OF GENERA AND SPECIES. -

FAO Fisheries & Aquaculture
Food and Agriculture Organization of the United Nations Fisheries and for a world without hunger Aquaculture Department Cultured Aquatic Species Information Programme Mercenaria mercenaria (Linnaeus, 1758) I. Identity V. Status And Trends a. Biological Features VI. Main Issues b. Images Gallery a. Responsible Aquaculture Practices II. Profile VII. References a. Historical Background a. Related Links b. Main Producer Countries c. Habitat And Biology III. Production a. Production Cycle b. Production Systems c. Diseases And Control Measures IV. Statistics a. Production Statistics b. Market And Trade Identity Mercenaria mercenaria Linnaeus, 1758 [Veneridae] FAO Names: En - Northern quahog(=Hard clam), Fr - Praire, Es - Chirla mercenaria Biological features Shell solid, equivalve; inequilateral, beaks in the front half of the shell; broadly oval in outline. Ligament a deeply inset, dark brown elliptical band, behind the beaks reaching half-way to the posterior margin. Lunule well defined, broad, heart-shaped. Escutcheon indistinct. Sculpture of concentric lines, raised here and there into ridges, and fine radiating lines. In young specimens the ridges are present all over the shell but in the adult they persist, after wear and tear, only near the anterior and posterior margins. Growth stages prominent. Both valves with three cardinal teeth; in addition there is present in each valve a rough tooth-like area behind the beaks and immediately below the ligament; this area has the appearance of a supplementary posterior cardinal tooth which has been broken off. No laterals. Pallial sinus not deep, triangular. Margin grenulate. Colour a dirty white, light varnish-brown, dull grey or grey-brown. Inside of shell white, sometimes deep violet about the adductor muscle scars. -

Transposed Hinge Structures in Lamellibranches
TRANSACTIONS OF THE • SAN DIEGO SOCIETY OF NATURAL HISTORY ,/ VoLUME VII, No. 26, pp. 299-318, plate 19 • TRANSPOSED HINGE STRUCTURES IN LAMELLIBRANCHS BY P. PoPENOE AND FINDLAY W. W. A. • Balch Graduate School of the Geological Sciences California Institute of Technology Pasadena, California SAN DIEGO, CALIFORNIA PRINTED FOR THE SOCIETY OCTOBER 6, 1933 • • COMMITTEE ON PUBLICATION U.S. GRANT, IV, Chairman FRED BAKER CLINTON G. ABBOTT, Editor • TRANSPOSED HINGE STRUCTURES IN LAMELLIBRANCHS 1 BY W. P. PoPENOE AND W. A. FINDLAY California Institute of Technology INTRODUCTION In the course of study of a collection of Eocene fossils from Claiborne Bluff, Alabama, the senior author of tl1is paper noticed two valves, one right and one left, of the lamellibranch Venericardia parva Lea, in whicl1 the dentition is partially transposed. Subsequently, the authors made an examination of more than five thousand lamellibranch valves, representit1g both recent and fossil shells, in search of further exa1nples of hinge-trans position. We have found a total number of twenty-six valves exhibiting this variation. Study of these specimens has revealed some hitherto unreported facts regarding the principles of hinge-transposition. Therefore in this paper, we shall describe and discuss these specimens, and shall present such con clusions as seem justified by the data assembled. Citations to the literature are made by author, date, and page, referring to the list at the end of the paper. DEFINITION OF T ERMS A transposed lamellibranch hinge is defined as one that exhibits in the right valve the hinge ele1nents normally occurring in the left valve, and vice-versa. -

71St Annual Meeting Society of Vertebrate Paleontology Paris Las Vegas Las Vegas, Nevada, USA November 2 – 5, 2011 SESSION CONCURRENT SESSION CONCURRENT
ISSN 1937-2809 online Journal of Supplement to the November 2011 Vertebrate Paleontology Vertebrate Society of Vertebrate Paleontology Society of Vertebrate 71st Annual Meeting Paleontology Society of Vertebrate Las Vegas Paris Nevada, USA Las Vegas, November 2 – 5, 2011 Program and Abstracts Society of Vertebrate Paleontology 71st Annual Meeting Program and Abstracts COMMITTEE MEETING ROOM POSTER SESSION/ CONCURRENT CONCURRENT SESSION EXHIBITS SESSION COMMITTEE MEETING ROOMS AUCTION EVENT REGISTRATION, CONCURRENT MERCHANDISE SESSION LOUNGE, EDUCATION & OUTREACH SPEAKER READY COMMITTEE MEETING POSTER SESSION ROOM ROOM SOCIETY OF VERTEBRATE PALEONTOLOGY ABSTRACTS OF PAPERS SEVENTY-FIRST ANNUAL MEETING PARIS LAS VEGAS HOTEL LAS VEGAS, NV, USA NOVEMBER 2–5, 2011 HOST COMMITTEE Stephen Rowland, Co-Chair; Aubrey Bonde, Co-Chair; Joshua Bonde; David Elliott; Lee Hall; Jerry Harris; Andrew Milner; Eric Roberts EXECUTIVE COMMITTEE Philip Currie, President; Blaire Van Valkenburgh, Past President; Catherine Forster, Vice President; Christopher Bell, Secretary; Ted Vlamis, Treasurer; Julia Clarke, Member at Large; Kristina Curry Rogers, Member at Large; Lars Werdelin, Member at Large SYMPOSIUM CONVENORS Roger B.J. Benson, Richard J. Butler, Nadia B. Fröbisch, Hans C.E. Larsson, Mark A. Loewen, Philip D. Mannion, Jim I. Mead, Eric M. Roberts, Scott D. Sampson, Eric D. Scott, Kathleen Springer PROGRAM COMMITTEE Jonathan Bloch, Co-Chair; Anjali Goswami, Co-Chair; Jason Anderson; Paul Barrett; Brian Beatty; Kerin Claeson; Kristina Curry Rogers; Ted Daeschler; David Evans; David Fox; Nadia B. Fröbisch; Christian Kammerer; Johannes Müller; Emily Rayfield; William Sanders; Bruce Shockey; Mary Silcox; Michelle Stocker; Rebecca Terry November 2011—PROGRAM AND ABSTRACTS 1 Members and Friends of the Society of Vertebrate Paleontology, The Host Committee cordially welcomes you to the 71st Annual Meeting of the Society of Vertebrate Paleontology in Las Vegas. -

Early Ontogeny of Jurassic Bakevelliids and Their Bearing on Bivalve Evolution
Early ontogeny of Jurassic bakevelliids and their bearing on bivalve evolution NIKOLAUS MALCHUS Malchus, N. 2004. Early ontogeny of Jurassic bakevelliids and their bearing on bivalve evolution. Acta Palaeontologica Polonica 49 (1): 85–110. Larval and earliest postlarval shells of Jurassic Bakevelliidae are described for the first time and some complementary data are given concerning larval shells of oysters and pinnids. Two new larval shell characters, a posterodorsal outlet and shell septum are described. The outlet is homologous to the posterodorsal notch of oysters and posterodorsal ridge of arcoids. It probably reflects the presence of the soft anatomical character post−anal tuft, which, among Pteriomorphia, was only known from oysters. A shell septum was so far only known from Cassianellidae, Lithiotidae, and the bakevelliid Kobayashites. A review of early ontogenetic shell characters strongly suggests a basal dichotomy within the Pterio− morphia separating taxa with opisthogyrate larval shells, such as most (or all?) Praecardioida, Pinnoida, Pterioida (Bakevelliidae, Cassianellidae, all living Pterioidea), and Ostreoida from all other groups. The Pinnidae appear to be closely related to the Pterioida, and the Bakevelliidae belong to the stem line of the Cassianellidae, Lithiotidae, Pterioidea, and Ostreoidea. The latter two superfamilies comprise a well constrained clade. These interpretations are con− sistent with recent phylogenetic hypotheses based on palaeontological and genetic (18S and 28S mtDNA) data. A more detailed phylogeny is hampered by the fact that many larval shell characters are rather ancient plesiomorphies. Key words: Bivalvia, Pteriomorphia, Bakevelliidae, larval shell, ontogeny, phylogeny. Nikolaus Malchus [[email protected]], Departamento de Geologia/Unitat Paleontologia, Universitat Autòno− ma Barcelona, 08193 Bellaterra (Cerdanyola del Vallès), Spain. -

The Marine and Brackish Water Mollusca of the State of Mississippi
Gulf and Caribbean Research Volume 1 Issue 1 January 1961 The Marine and Brackish Water Mollusca of the State of Mississippi Donald R. Moore Gulf Coast Research Laboratory Follow this and additional works at: https://aquila.usm.edu/gcr Recommended Citation Moore, D. R. 1961. The Marine and Brackish Water Mollusca of the State of Mississippi. Gulf Research Reports 1 (1): 1-58. Retrieved from https://aquila.usm.edu/gcr/vol1/iss1/1 DOI: https://doi.org/10.18785/grr.0101.01 This Article is brought to you for free and open access by The Aquila Digital Community. It has been accepted for inclusion in Gulf and Caribbean Research by an authorized editor of The Aquila Digital Community. For more information, please contact [email protected]. Gulf Research Reports Volume 1, Number 1 Ocean Springs, Mississippi April, 1961 A JOURNAL DEVOTED PRIMARILY TO PUBLICATION OF THE DATA OF THE MARINE SCIENCES, CHIEFLY OF THE GULF OF MEXICO AND ADJACENT WATERS. GORDON GUNTER, Editor Published by the GULF COAST RESEARCH LABORATORY Ocean Springs, Mississippi SHAUGHNESSY PRINTING CO.. EILOXI, MISS. 0 U c x 41 f 4 21 3 a THE MARINE AND BRACKISH WATER MOLLUSCA of the STATE OF MISSISSIPPI Donald R. Moore GULF COAST RESEARCH LABORATORY and DEPARTMENT OF BIOLOGY, MISSISSIPPI SOUTHERN COLLEGE I -1- TABLE OF CONTENTS Introduction ............................................... Page 3 Historical Account ........................................ Page 3 Procedure of Work ....................................... Page 4 Description of the Mississippi Coast ....................... Page 5 The Physical Environment ................................ Page '7 List of Mississippi Marine and Brackish Water Mollusca . Page 11 Discussion of Species ...................................... Page 17 Supplementary Note ..................................... -

Molluscs (Mollusca: Gastropoda, Bivalvia, Polyplacophora)
Gulf of Mexico Science Volume 34 Article 4 Number 1 Number 1/2 (Combined Issue) 2018 Molluscs (Mollusca: Gastropoda, Bivalvia, Polyplacophora) of Laguna Madre, Tamaulipas, Mexico: Spatial and Temporal Distribution Martha Reguero Universidad Nacional Autónoma de México Andrea Raz-Guzmán Universidad Nacional Autónoma de México DOI: 10.18785/goms.3401.04 Follow this and additional works at: https://aquila.usm.edu/goms Recommended Citation Reguero, M. and A. Raz-Guzmán. 2018. Molluscs (Mollusca: Gastropoda, Bivalvia, Polyplacophora) of Laguna Madre, Tamaulipas, Mexico: Spatial and Temporal Distribution. Gulf of Mexico Science 34 (1). Retrieved from https://aquila.usm.edu/goms/vol34/iss1/4 This Article is brought to you for free and open access by The Aquila Digital Community. It has been accepted for inclusion in Gulf of Mexico Science by an authorized editor of The Aquila Digital Community. For more information, please contact [email protected]. Reguero and Raz-Guzmán: Molluscs (Mollusca: Gastropoda, Bivalvia, Polyplacophora) of Lagu Gulf of Mexico Science, 2018(1), pp. 32–55 Molluscs (Mollusca: Gastropoda, Bivalvia, Polyplacophora) of Laguna Madre, Tamaulipas, Mexico: Spatial and Temporal Distribution MARTHA REGUERO AND ANDREA RAZ-GUZMA´ N Molluscs were collected in Laguna Madre from seagrass beds, macroalgae, and bare substrates with a Renfro beam net and an otter trawl. The species list includes 96 species and 48 families. Six species are dominant (Bittiolum varium, Costoanachis semiplicata, Brachidontes exustus, Crassostrea virginica, Chione cancellata, and Mulinia lateralis) and 25 are commercially important (e.g., Strombus alatus, Busycoarctum coarctatum, Triplofusus giganteus, Anadara transversa, Noetia ponderosa, Brachidontes exustus, Crassostrea virginica, Argopecten irradians, Argopecten gibbus, Chione cancellata, Mercenaria campechiensis, and Rangia flexuosa). -

Civitanova Marche
ISPRA Istituto Superiore per la Protezione e la Ricerca Ambientale SERVIZIO GEOLOGICO D’ITALIA Organo Cartografico dello Stato (legge n° 68 del 2.2.1960) NOTE ILLUSTRATIVE della CARTA GEOLOGICA D’ITALIA alla scala 1:50.000 foglio 304 CIVITANOVA MARCHE A cura di G. Cantalamessa(1) Con contributi di: Sedimentologia: C. Di Celma(1), M. Piccini(1) Stratigrafi a: P. Didaskalou(1) , M. Potetti(1), P. Lori(1) Area marina: Interpretazione: E. Campiani(2), A. Correggiari(2), F. Foglini(2), F. Trincardi(2) Micropaleontologia: A. Asioli(3) PROGETTOSedimentologia e Radionuclidi: A. Gallerani(2), L. Langone(2), F. Lorenzini(2) (1) Dipartimento di Scienze della Terra, Università di Camerino (2) CNR-ISMAR, Bologna (3) CNR-IGG, Padova Ente realizzatore: nnote_304_11_2011.inddote_304_11_2011.indd 1 CARG117/11/117/11/11 111.291.29 Direttore del Servizio Geologico d’Italia - ISPRA: C. CAMPOBASSO Responsabile del Progetto CARG per il Servizio Geologico d’Italia - ISPRA: F. GALLUZZO Responsabile del Progetto CARG per Regione Marche: M. PRINCIPI PER IL SERVIZIO GEOLOGICO D’ITALIA - ISPRA: Revisione scientifi ca: F. Capotorti, D. Delogu, C. Muraro, S. Nisio S. D’Angelo, A. Fiorentino (parte a mare) Coordinamento cartografi co: D. Tacchia (coord.), V. Pannuti Revisione informatizzata dei dati geologici: L. Battaglini, V. Campo; M. Rossi (ASC) Coordinamento editoriale e allestimento per la stampa: D. Tacchia, V. Pannuti PER LA REGIONE MARCHE: Allestimento editoriale e cartografi co: M. Piccini(1), P. Didaskalou(1), P. Lori (1) (1) Dipartimento di Scienze della Terra, Università di Camerino Allestimento editoriale e cartografi co (area marina): E. Campiani Allestimento informatizzazione dei dati geologici (area marina): F.