What Is the Function of the Cone-Rich Ri M of the Retina?
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Meet the Choroid N Optometric Management Joe Pizzimenti, OD, FAAO O Scientific Advisory Boards [email protected] N Zeiss N Zeavision N Thrombogenics N Genentech
10/29/19 Financial Disclosures o Honoraria n Review of Optometry Meet The Choroid n Optometric Management Joe Pizzimenti, OD, FAAO o Scientific Advisory Boards [email protected] n Zeiss n Zeavision n Thrombogenics n Genentech Financial Disclosures Goals for This Course o Consulting Fees n Zeiss o Functional anatomy review n Zeavision n Choroid n Maculogix o Choroid examination and evaluation o Proprietary Interests o Case examples n None o Interactive o Stockholder: Zeavision Questions? 1 10/29/19 The Choroid The Choroid: Structure, o Located between the Function, and Evaluation sclera and the RPE n Extends from ora serrata to optic nerve o Pigmented/vascular tissue .75mm thick o Nourishes the RPE n Choroiocapillaris designed to leak o Absorbs light that passes through retina The Choriod RPE Bruch’s Membrane thickness o Loose connective tissue o Basal lamina of RPE o Melanocytes o Anterior collagenous o Choriocapillaris layer Mel. n Fenestrated endothelium o Elastic layer allows diffusion of o Posterior collagenous proteins CC layer n S__________ regulation o Basal lamina of CC BM n High blood flow endothelium n Very little O-2 extracted, o Contamination of so high venous O-2 Bruch’s can result in sclera d________, CNVM Nourishing the Retina Choroid Microstructure o 2 main sources of blood supply to retina: • Choriocapillaris o Choroidal BVs n Supplies outer retinal • Sattler’s layer layers, including PRs o CRA • Haller’s layer n 4 branches nourish inner retina • Supra - choroid n Run radially toward fovea 2 10/29/19 Imaging the Vascular Layers Imaging the Choroid of the Choroid WHAT IS ENHANCED Imaging the Choroid-EDI DEPTH OCT IMAGING? • EDI-OCT • Enhanced-depth imaging (EDI) OCT modifies the standard technique of image acquisition to better reveal the structural details of the choroid. -

Torpedo Maculopathy at the Site of the Fetal “Bulge”
SMALL CASE SERIES SECTION EDITOR: W. RICHARD GREEN, MD ward the foveola. This defect closely The flat, nonpigmented lesion mea- Torpedo Maculopathy resembles solitary CHRPE but dif- sured 2 mm horizontally and 1 mm at the Site of the fers in its nonrandom macular lo- vertically and was located 4 mm tem- Fetal “Bulge” cation and pointed torpedo shape.5-8 poral to the optic disc (Figure 2). In the few reported cases, there have Toxoplasmosis titer results were been no systemic associations. negative. Observation was advised. Torpedo maculopathy was discov- Herein, we describe 2 cases of tor- ered in 2 children as a pointed-oval pedo maculopathy and speculate as Comment. In 1992, Roseman and retinal pigment epithelial (RPE) de- to its embryogenesis. Gass3 described a 12-year-old boy fect in the temporal macula. This with a small, flat, circumscribed, oval congenital finding could be related Report of Cases. Case 1. On rou- RPE lesion in the temporal macula. to the fetal temporal macular “bulge” tine eye examination, a 3-year-old Additional reports confirmed the con- that normally occurs at 4 to 6 girl with fix-and-follow visual acu- sistent pointed oval configuration and months’ gestation at the same site. ity was discovered to have a tempo- macular location of this condition There are several congenital ral macular RPE defect with a (Table).5-8 Rigotti and associates7 re- anomalies of the RPE, including con- pointed-oval shape directed toward ported 3 cases of asymptomatic tor- genital hypertrophy of the RPE the foveola and hyperpigmented pedo maculopathy in a child and 2 (CHRPE), combined hamartoma of “frayed tail” appearance directed to- adults. -

Ciliary Body
Ciliary body S.Karmakar HOD Introduction • Ciliary body is the middle part of the uveal tract . It is a ring (slightly eccentric ) shaped structure which projects posteriorly from the scleral spur, with a meridional width varying from 5.5 to 6.5 mm. • It is brown in colour due to melanin pigment. Anteriorly it is confluent with the periphery of the iris (iris root) and anterior part of the ciliary body bounds a part of the anterior chamber angle. Introduction • Posteriorly ciliary body has a crenated or scalloped periphery, known as ora serrata, where it is continuous with the choroid and retina. The ora serrata exhibits forward extensions,known as dentate process, which are well defined on the nasal side and less so temporally. • Ciliary body has a width of approximately 5.9 mm on the nasal side and 6.7 mm on the temporal side. Extension of the ciliary body On the outside of the eyeball, the ciliary body extends from a point about 1.5 mm posterior to the corneal limbus to a point 6.5 to 7.5 mm posterior to this point on the temporal side and 6.5 mm posterior on the nasal side. Parts of ciliary body • Ciliary body, in cross section, is a triangular structure ( in diagram it can be compared as ∆ AOI). Outer side of the triangle (O) is attached with the sclera with suprachoroidal space in between. Anterior side of the triangle (A) forms part of the anterior & posterior chamber. In its middle, the iris is attached. The inner side of the triangle (I) is divided into two parts. -

Nomina Histologica Veterinaria, First Edition
NOMINA HISTOLOGICA VETERINARIA Submitted by the International Committee on Veterinary Histological Nomenclature (ICVHN) to the World Association of Veterinary Anatomists Published on the website of the World Association of Veterinary Anatomists www.wava-amav.org 2017 CONTENTS Introduction i Principles of term construction in N.H.V. iii Cytologia – Cytology 1 Textus epithelialis – Epithelial tissue 10 Textus connectivus – Connective tissue 13 Sanguis et Lympha – Blood and Lymph 17 Textus muscularis – Muscle tissue 19 Textus nervosus – Nerve tissue 20 Splanchnologia – Viscera 23 Systema digestorium – Digestive system 24 Systema respiratorium – Respiratory system 32 Systema urinarium – Urinary system 35 Organa genitalia masculina – Male genital system 38 Organa genitalia feminina – Female genital system 42 Systema endocrinum – Endocrine system 45 Systema cardiovasculare et lymphaticum [Angiologia] – Cardiovascular and lymphatic system 47 Systema nervosum – Nervous system 52 Receptores sensorii et Organa sensuum – Sensory receptors and Sense organs 58 Integumentum – Integument 64 INTRODUCTION The preparations leading to the publication of the present first edition of the Nomina Histologica Veterinaria has a long history spanning more than 50 years. Under the auspices of the World Association of Veterinary Anatomists (W.A.V.A.), the International Committee on Veterinary Anatomical Nomenclature (I.C.V.A.N.) appointed in Giessen, 1965, a Subcommittee on Histology and Embryology which started a working relation with the Subcommittee on Histology of the former International Anatomical Nomenclature Committee. In Mexico City, 1971, this Subcommittee presented a document entitled Nomina Histologica Veterinaria: A Working Draft as a basis for the continued work of the newly-appointed Subcommittee on Histological Nomenclature. This resulted in the editing of the Nomina Histologica Veterinaria: A Working Draft II (Toulouse, 1974), followed by preparations for publication of a Nomina Histologica Veterinaria. -
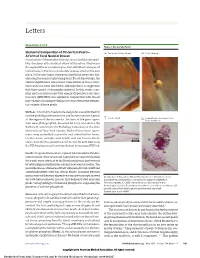
Elemental Composition of Ora Serrata Pearls—A Form of Focal Nodular
Letters RESEARCH LETTER Figure 1. Ora Serrata Pearls Elemental Composition of Ora Serrata Pearls— A Two pearls (50 and 10 µm) B Detailed image A Form of Focal Nodular Drusen Examination of the peripheral fundus may reveal discrete pearl- like structures at the furthest extent of the retina. They occur throughout the ora serrata region, but only where a tongue of retina tissue in the form of a dentate process overlies the pars plana. In the later stages, the pigment epithelial covering is lost, exposing the pearl as a glistening bead. To our knowledge, the clinical significance and precise composition of these struc- tures have not been described, although there is suggestion that they consist of drusenlike material. In this study, scan- ning electron microscopy with energy dispersive x-ray spec- troscopy (SEM-EDS) was applied in conjunction with classic histochemical staining techniques to characterize the elemen- tal content of these pearls. Methods | A formalin-fixed enucleated globe was submitted for routine pathological examination and found to contain 3 pearls C Calcified pearl D Pearl without attachment to the at the region of the ora serrata. Sections of the gross speci- Bruch membrane men were photographed, dissected (R.E.E.), and sent to the Barbara W. Streeten Ocular Pathology Laboratory at the State University of New York Upstate Medical University. Speci- mens were embedded in paraffin and submitted for hema- toxylin-eosin, periodic acid–Schiff, and van Gieson elastic 200 µm stains. A section was placed on a carbon stub for SEM-EDS using the FEI Aspex personal scanning electron microscope (FEI Inc). -

The Horizontal Raphe of the Human Retina and Its Watershed Zones
vision Review The Horizontal Raphe of the Human Retina and its Watershed Zones Christian Albrecht May * and Paul Rutkowski Department of Anatomy, Medical Faculty Carl Gustav Carus, TU Dresden, 74, 01307 Dresden, Germany; [email protected] * Correspondence: [email protected] Received: 24 September 2019; Accepted: 6 November 2019; Published: 8 November 2019 Abstract: The horizontal raphe (HR) as a demarcation line dividing the retina and choroid into separate vascular hemispheres is well established, but its development has never been discussed in the context of new findings of the last decades. Although factors for axon guidance are established (e.g., slit-robo pathway, ephrin-protein-receptor pathway) they do not explain HR formation. Early morphological organization, too, fails to establish a HR. The development of the HR is most likely induced by the long posterior ciliary arteries which form a horizontal line prior to retinal organization. The maintenance might then be supported by several biochemical factors. The circulation separate superior and inferior vascular hemispheres communicates across the HR only through their anastomosing capillary beds resulting in watershed zones on either side of the HR. Visual field changes along the HR could clearly be demonstrated in vascular occlusive diseases affecting the optic nerve head, the retina or the choroid. The watershed zone of the HR is ideally protective for central visual acuity in vascular occlusive diseases but can lead to distinct pathological features. Keywords: anatomy; choroid; development; human; retina; vasculature 1. Introduction The horizontal raphe (HR) was first described in the early 1800s as a horizontal demarcation line that extends from the macula to the temporal Ora dividing the temporal retinal nerve fiber layer into a superior and inferior half [1]. -

The Nervous System: General and Special Senses
18 The Nervous System: General and Special Senses PowerPoint® Lecture Presentations prepared by Steven Bassett Southeast Community College Lincoln, Nebraska © 2012 Pearson Education, Inc. Introduction • Sensory information arrives at the CNS • Information is “picked up” by sensory receptors • Sensory receptors are the interface between the nervous system and the internal and external environment • General senses • Refers to temperature, pain, touch, pressure, vibration, and proprioception • Special senses • Refers to smell, taste, balance, hearing, and vision © 2012 Pearson Education, Inc. Receptors • Receptors and Receptive Fields • Free nerve endings are the simplest receptors • These respond to a variety of stimuli • Receptors of the retina (for example) are very specific and only respond to light • Receptive fields • Large receptive fields have receptors spread far apart, which makes it difficult to localize a stimulus • Small receptive fields have receptors close together, which makes it easy to localize a stimulus. © 2012 Pearson Education, Inc. Figure 18.1 Receptors and Receptive Fields Receptive Receptive field 1 field 2 Receptive fields © 2012 Pearson Education, Inc. Receptors • Interpretation of Sensory Information • Information is relayed from the receptor to a specific neuron in the CNS • The connection between a receptor and a neuron is called a labeled line • Each labeled line transmits its own specific sensation © 2012 Pearson Education, Inc. Interpretation of Sensory Information • Classification of Receptors • Tonic receptors -

Anatomy of the Globe 09 Hermann D. Schubert Basic and Clinical
Anatomy of the Globe 09 Hermann D. Schubert Basic and Clinical Science Course, AAO 2008-2009, Section 2, Chapter 2, pp 43-92. The globe is the home of the retina (part of the embryonic forebrain, i.e.neural ectoderm and neural crest) which it protects, nourishes, moves or holds in proper position. The retinal ganglion cells (second neurons of the visual pathway) have axons which form the optic nerve (a brain tract) and which connect to the lateral geniculate body of the brain (third neurons of the visual pathway with axons to cerebral cortex). The transparent media of the eye are: tear film, cornea, aqueous, lens, vitreous, internal limiting membrane and inner retina. Intraocular pressure is the pressure of the aqueous and vitreous compartment. The aqueous compartment is comprised of anterior(200ul) and posterior chamber(60ul). Aqueous and vitreous compartments communicate across the anterior cortical gel of the vitreous which seen from up front looks like a donut and is called the “annular diffusional gap.” The globe consists of two superimposed spheres, the corneal radius measuring 8mm and the scleral radius 12mm. The superimposition creates an external scleral sulcus, the outflow channels anterior to the scleral spur fill the internal scleral sulcus. Three layers or ocular coats are distinguished: the corneal scleral coat, the uvea and neural retina consisting of retina and pigmentedepithelium. The coats and components of the inner eye are held in place by intraocular pressure, scleral rigidity and mechanical attachments between the layers. The corneoscleral coat consists of cornea, sclera, lamina cribrosa and optic nerve sheath. -

Nervous and Endocrine Systems Lab Guide
CSM Human Anatomy Nervous and Endocrine Systems Lab Guide Histology White matter Spinal cord (16) features Cerebrum including: Convolutions - gyri motor neuron in anterior gray Sulci horn Cerebral cortex sensory/dorsal root ganglion R & L hemispheres with cell bodies of sensory longitudinal fissure nerves frontal lobe central canal central sulcus posterior(dorsal) horns lateral sulcus anterior(ventral) horns parieto-occipital lateral horns sulcus gray commissure precentral gyrus dorsal root ganglia Broca's area nerve roots parietal lobe retina, choroid, sclera (17) postcentral gyrus Organ of Corti (18) temporal lobe thyroid follicles (34) occipital lobe adrenal gland - medulla and insula cortex (36) basal nuclei pituitary gland - anterior and transverse fissure posterior (35) corpus callosum, corona pancreas – pancreatic islets (of radiata, internal capsule Langerhans) (25) mesencephalon corpora quadrigemina Central Nervous System superior colliculus Brain inferior colliculus Meninges substantia nigra dura mater cerebellum falx cerebri arbor vitae falx cerebelli cerebellar cortex tentorium cerebelli thalamus dural venous sinus intermediate pia mater mass/interthalamic arachnoid membrane adhesion subarachnoid space hypothalamus arachnoid villi pineal body choroid plexus pituitary gland Gray matter infundibulum 1 pons Sympathetic Chain of Ganglia medulla oblongata lateral ventricles Eye third ventricle fibrous tunic cerebral aqueduct sclera fourth ventricle cornea Spinal Cord vascular tunic central canal choroid conus medullaris ciliary -

Peripheral Lesions of the Fundus
Peripheral Lesions of the Fundus David W. Faber, M.D. Financial Disclosure The author has no financial interest in the subject or in any products discussed. Examination Techniques Slit Lamp Biomiroscopy Indirect Ophthalmoscopy With and without scleral depression Wide Angle Photography and Angiography OCT (Limited value) Normal Anatomy Pars Plana Ora Serrata Vitreous Base Peripheral Retinal Normal Fundus Pars Plana Ora Serrata Ora Serrata Lesions of the Ora Serrata and Pars Plana Meridional Folds Meridional Complex Fold that extends to ciliary process Pars Plana Cysts Ora Serrata Pearls Meridional Fold Meridional Fold with Tear Meridional Complex (Fold) Enclosed Oral Bay Pars Plana Cysts Ora Serrata Pearl Diffuse Areas of Whitening White with and without Pressure Snowbanking Retinitis White Without Pressure White With Pressure Snowbanking CMV Retinitis Acute Retinal Necrosis Focal Areas of Change Peripheral Cystoid Degeneration Cobblestone Degeneration Equatorial Drusen Fleck Retina Syndromes Retinal Tufts Coloboma Retinal holes and tears Peripheral Cystoid Degeneration Peripheral Cystoid Degeneration Pavingstone Degeneration Pavingstone Degeneration Cuticular Drusen Fundus Albipunctatus Retinitis Punctata Albescens Fundus Flavimaculata Stargardt's Disease Retinal Tuft Chorioretinal Coloboma Horseshoe Tear Horseshoe Tear s/p Laser Operculated Retinal hole Diffuse Pigmentary changes Lattice Degeneration Peripheral Reticular Pigment Degeneration (PRPD) Retinitis Pigmentosa Gyrate Atrophy Diffuse Pigmentary changes Inflammatory/Infectious -

Eye Anatomy Slides File
EYE ANATOMY & PHYSIOLOGY Dr. Cesar Carrillo [email protected] Sight For All **Disclaimer** The images contained in this presentaon are not my own, they can be found on the web Eye Anatomy and Physiology A thorough understanding of the anatomy and physiology of the eye, orbit, visual pathways, upper cranial nerves, and central pathways for the control of eye movements is a prerequisite for proper interpretaon of diseases having ocular manifestaons. Furthermore, such anatomic knowledge is essen;al to the proper planning and safe execu;on of ocular and orbital surgery Eye Anatomy and Physiology Objecves: Brief overview of eye anatomy and relevant physiology — Embriology — The Orbit, Cranial Nerves, Blood supply and Venous drainage — The Ocular Adnexa — The Extraocular muscles — The Conjunc;va, Sclera and Cornea — The Uveal tract — The Lens — The Re;na, Vitreous and Op;c Nerve — The Visual Pathway Embryology The eye is derived from three of the primi;ve embryonic layers: — Surface ectoderm, including its derivave the neural crest — Neural ectoderm — Mesoderm — Endoderm does not enter into the formaon of the eye — Mesenchyme is the term for embryonic connecve ssue. Ocular and adnexal connec;ve ;ssues previously were thought to be derived from mesoderm, but it has now been shown that most of the mesenchyme of all of the head and neck region is derived from the cranial neural crest Embryology Development of the structures of the head and neck occurs between 3-8 weeks of gestation — Eye develops as an ectodermal diverticulum from the lateral -

26 April 2010 TE Prepublication Page 1 Nomina Generalia General Terms
26 April 2010 TE PrePublication Page 1 Nomina generalia General terms E1.0.0.0.0.0.1 Modus reproductionis Reproductive mode E1.0.0.0.0.0.2 Reproductio sexualis Sexual reproduction E1.0.0.0.0.0.3 Viviparitas Viviparity E1.0.0.0.0.0.4 Heterogamia Heterogamy E1.0.0.0.0.0.5 Endogamia Endogamy E1.0.0.0.0.0.6 Sequentia reproductionis Reproductive sequence E1.0.0.0.0.0.7 Ovulatio Ovulation E1.0.0.0.0.0.8 Erectio Erection E1.0.0.0.0.0.9 Coitus Coitus; Sexual intercourse E1.0.0.0.0.0.10 Ejaculatio1 Ejaculation E1.0.0.0.0.0.11 Emissio Emission E1.0.0.0.0.0.12 Ejaculatio vera Ejaculation proper E1.0.0.0.0.0.13 Semen Semen; Ejaculate E1.0.0.0.0.0.14 Inseminatio Insemination E1.0.0.0.0.0.15 Fertilisatio Fertilization E1.0.0.0.0.0.16 Fecundatio Fecundation; Impregnation E1.0.0.0.0.0.17 Superfecundatio Superfecundation E1.0.0.0.0.0.18 Superimpregnatio Superimpregnation E1.0.0.0.0.0.19 Superfetatio Superfetation E1.0.0.0.0.0.20 Ontogenesis Ontogeny E1.0.0.0.0.0.21 Ontogenesis praenatalis Prenatal ontogeny E1.0.0.0.0.0.22 Tempus praenatale; Tempus gestationis Prenatal period; Gestation period E1.0.0.0.0.0.23 Vita praenatalis Prenatal life E1.0.0.0.0.0.24 Vita intrauterina Intra-uterine life E1.0.0.0.0.0.25 Embryogenesis2 Embryogenesis; Embryogeny E1.0.0.0.0.0.26 Fetogenesis3 Fetogenesis E1.0.0.0.0.0.27 Tempus natale Birth period E1.0.0.0.0.0.28 Ontogenesis postnatalis Postnatal ontogeny E1.0.0.0.0.0.29 Vita postnatalis Postnatal life E1.0.1.0.0.0.1 Mensurae embryonicae et fetales4 Embryonic and fetal measurements E1.0.1.0.0.0.2 Aetas a fecundatione5 Fertilization