Functional Ecological Convergence Between the Thylacine and Small Prey-Focused Canids
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Fur Trade and the Biotic Homogenization of Subpolar Ecosystems
Chapter 14 Fur Trade and the Biotic Homogenization of Subpolar Ecosystems Ramiro D. Crego, Ricardo Rozzi, and Jaime E. Jiménez Abstract At the southern end of the Americas exist one of the last pristine ecosys- tems in the world, the sub-Antarctic Magellanic forests ecoregion, protected by the Cape Horn Biosphere Reserve (CHBR). Despite its remote location, the CHBR has been subject to the growing infuences of globalization, a process that has driven cultural, biotic, and economic transformations in the region since the mid-twentieth century. One of the most important threats to these unique ecosystems is the increase of biological invasions. Motivated by the expanding fur industry that responded to the globalization process, American beavers (Castor canadensis), muskrats (Ondatra zibethicus), and American minks (Neovison vison) were introduced, inde- pendently, to the southern tip of South America. Research has shown that these three North American species have reassembled their native interactions to affect negatively the invaded ecosystems of the CHBR. Beavers affect river fow and R. D. Crego (*) Department of Biological Sciences, University of North Texas, Denton, TX, USA Instituto de Ecología y Biodiversidad, Santiago, Chile Sub-Antarctic Biocultural Conservation Program, University of North Texas, Denton, TX, USA R. Rozzi Department of Philosophy and Religion and Department of Biological Sciences, University of North Texas, Denton, TX, USA Sub-Antarctic Biocultural Conservation Program, University of North Texas, Denton, TX, USA Instituto de Ecología y Biodiversidad and Universidad de Magallanes, Punta Arenas, Chile J. E. Jiménez Department of Biological Sciences, University of North Texas, Denton, TX, USA Instituto de Ecología y Biodiversidad, Santiago, Chile Sub-Antarctic Biocultural Conservation Program, University of North Texas, Denton, TX, USA Department of Philosophy and Religion, University of North Texas, Denton, TX, USA Universidad de Magallanes, Punta Arenas, Chile © Springer Nature Switzerland AG 2018 233 R. -
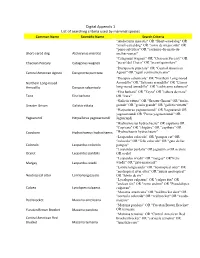
Digital Appendix 1 List of Searching Criteria Used by Mammal Species
Digital Appendix 1 List of searching criteria used by mammal species Common Name Scientific Name Search Criteria "Atelocynus microtis" OR "Short-eared dog" OR "small-eared dog" OR "zorro de oreja corta" OR "perro selvático" OR "cachorro-do-mato-de- Short-eared dog Atelocynus microtis orelhas-curtas" "Catagonus wagneri" OR "Chacoan Peccary" OR Chacoan Peccary Catagonus wagneri "pecarí del Chaco" OR "pecarí quimilero" “Dasyprocta punctate” OR "Central American Central American Agouti Dasyprocta punctata Agouti" OR "agutí centroamericano" “Dasypus sabanicola” OR "Northern Long-nosed Northern Long-nosed Armadillo" OR "Savanna armadillo" OR "Llanos Armadillo Dasypus sabanicola long-nosed armadillo" OR "cachicamo sabanero" “Eira barbara” OR "Tayra" OR "cabeza de mate" Taira Eira barbara OR "irara" “Galictis vittata” OR "Greater Grison" OR "furão- Greater Grison Galictis vittata grande" OR "grisón grande" OR "galictis vittatta" “Herpailurus yagouaroundi” OR Yaguarundi OR yagouaroundi OR "Puma yagouaroundi" OR Yaguarundi Herpailurus yagouaroundi Jaguarundi "Hydrochoerus hydrochaeris" OR capybara OR "Capivara" OR "chigüire" OR "capibara" OR Capybara Hydrochoerus hydrochaeris "Hydrochaeris hydrochaeris" “Leopardus colocolo” OR "pampas cat" OR "colocolo" OR "felis colocolo" OR "gato de las Colocolo Leopardus colocolo pampas" "Leopardus pardalis" OR jaguatirica OR ocelote Ocelot Leopardus pardalis OR ocelot “Leopardus wiedii” OR "margay" OR"Felis Margay Leopardus wiedii wiedii" OR "gato-maracajá" “Lontra longicaudis” OR "neotropical otter" OR "neotropical -

The Hunter and Biodiversity in Tasmania
The Hunter and Biodiversity in Tasmania The Hunter takes place on Tasmania’s Central Plateau, where “One hundred and sixty-five million years ago potent forces had exploded, clashed, pushed the plateau hundreds of metres into the sky.” [a, 14] The story is about the hunt for the last Tasmanian tiger, described in the novel as: “that monster whose fabulous jaw gapes 120 degrees, the carnivorous marsupial which had so confused the early explorers — a ‘striped wolf’, ‘marsupial wolf.’” [a, 16] Fig 1. Paperbark woodlands and button grass plains near Derwent Bridge, Central Tasmania. Source: J. Stadler, 2010. Biodiversity “Biodiversity”, or biological diversity, refers to variety in all forms of life—all plants and animals, their genes, and the ecosystems they live in. [b] It is important because all living things are connected with each other. For example, humans depend on living things in the environment for clean air to breathe, food to eat, and clean water to drink. Biodiversity is one of the underlying themes in The Hunter, a Tasmanian film directed by David Nettheim in 2011 and based on Julia Leigh’s 1999 novel about the hunt for the last Tasmanian Tiger. The film and the novel showcase problems that arise from loss of species, loss of habitat, and contested ideas about land use. The story is set in the Central Plateau Conservation Area and much of the film is shot just south of that area near Derwent Bridge and in the Florentine Valley. In Tasmania, land clearing is widely considered to be the biggest threat to biodiversity [c, d]. -

Thylacinidae
FAUNA of AUSTRALIA 20. THYLACINIDAE JOAN M. DIXON 1 Thylacine–Thylacinus cynocephalus [F. Knight/ANPWS] 20. THYLACINIDAE DEFINITION AND GENERAL DESCRIPTION The single member of the family Thylacinidae, Thylacinus cynocephalus, known as the Thylacine, Tasmanian Tiger or Wolf, is a large carnivorous marsupial (Fig. 20.1). Generally sandy yellow in colour, it has 15 to 20 distinct transverse dark stripes across the back from shoulders to tail. While the large head is reminiscent of the dog and wolf, the tail is long and characteristically stiff and the legs are relatively short. Body hair is dense, short and soft, up to 15 mm in length. Body proportions are similar to those of the Tasmanian Devil, Sarcophilus harrisii, the Eastern Quoll, Dasyurus viverrinus and the Tiger Quoll, Dasyurus maculatus. The Thylacine is digitigrade. There are five digital pads on the forefoot and four on the hind foot. Figure 20.1 Thylacine, side view of the whole animal. (© ABRS)[D. Kirshner] The face is fox-like in young animals, wolf- or dog-like in adults. Hairs on the cheeks, above the eyes and base of the ears are whitish-brown. Facial vibrissae are relatively shorter, finer and fewer than in Tasmanian Devils and Quolls. The short ears are about 80 mm long, erect, rounded and covered with short fur. Sexual dimorphism occurs, adult males being larger on average. Jaws are long and powerful and the teeth number 46. In the vertebral column there are only two sacrals instead of the usual three and from 23 to 25 caudal vertebrae rather than 20 to 21. -

The Impact of Forest Logging and Fragmentation on Carnivore Species Composition, Density and Occupancy in Madagascar’S Rainforests
The impact of forest logging and fragmentation on carnivore species composition, density and occupancy in Madagascar’s rainforests B RIAN D. GERBER,SARAH M. KARPANTY and J OHNY R ANDRIANANTENAINA Abstract Forest carnivores are threatened globally by Introduction logging and forest fragmentation yet we know relatively little about how such change affects predator populations. arnivores are one of the most threatened groups of 2009 This is especially true in Madagascar, where carnivores Cterrestrial mammals (Karanth & Chellam, ). have not been extensively studied. To understand better the Declines of predators are often attributed to habitat loss effects of logging and fragmentation on Malagasy carnivores and fragmentation but few quantitative studies have we evaluated species composition, density of fossa examined how carnivore populations and communities 2002 Cryptoprocta ferox and Malagasy civet Fossa fossana, and change with habitat loss or fragmentation (Crooks, ; 2005 carnivore occupancy in central-eastern Madagascar. We Michalski & Peres, ). This is particularly true for ’ photographically-sampled carnivores in two contiguous Madagascar s carnivores, with knowledge lacking about ff (primary and selectively-logged) and two fragmented rain- their ecology and the e ects of anthropogenic disturbances 2010 forests (fragments , 2.5 and . 15 km from intact forest). (Irwin et al., ), especially in the eastern rainforest where Species composition varied, with more native carnivores in only short-term studies have been conducted (Gerber et al., 2010 16 the contiguous than fragmented rainforests. F. fossana was ). With only % of the original primary forests extant absent from fragmented rainforests and at a lower density in Madagascar and those remaining becoming smaller and 2007 in selectively-logged than in primary rainforest (mean more isolated over time (Harper et al., ), habitat loss −2 1.38 ± SE 0.22 and 3.19 ± SE 0.55 individuals km , respect- and fragmentation are serious threats to many endemic 2010 ively). -

Redalyc.TRENDS in RESEARCH on TERRESTRIAL SPECIES of THE
Mastozoología Neotropical ISSN: 0327-9383 [email protected] Sociedad Argentina para el Estudio de los Mamíferos Argentina Pérez-Irineo, Gabriela; Santos-Moreno, Antonio TRENDS IN RESEARCH ON TERRESTRIAL SPECIES OF THE ORDER CARNIVORA Mastozoología Neotropical, vol. 20, núm. 1, 2013, pp. 113-121 Sociedad Argentina para el Estudio de los Mamíferos Tucumán, Argentina Available in: http://www.redalyc.org/articulo.oa?id=45728549008 How to cite Complete issue Scientific Information System More information about this article Network of Scientific Journals from Latin America, the Caribbean, Spain and Portugal Journal's homepage in redalyc.org Non-profit academic project, developed under the open access initiative Mastozoología Neotropical, 20(1):113-121, Mendoza, 2013 Copyright ©SAREM, 2013 Versión impresa ISSN 0327-9383 http://www.sarem.org.ar Versión on-line ISSN 1666-0536 Artículo TRENDS IN RESEARCH ON TERRESTRIAL SPECIES OF THE ORDER CARNIVORA Gabriela Pérez-Irineo and Antonio Santos-Moreno Laboratorio de Ecología Animal, Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional, Unidad Oaxaca, Instituto Politécnico Nacional, Hornos 1003, 71230 Santa Cruz Xoxocotlán, Oaxaca, México [Correspondence: Gabriela Pérez Irineo <[email protected]>]. ABSTRACT. Information regarding trends in research on terrestrial species of the order Carnivora can provide an understanding of the degree of knowledge of the order, or lack thereof, as well as help identifying areas on which to focus future research efforts. With the aim of providing information on these trends, this work presents a review of the thematic focuses of studies addressing this order published over the past three de- cades. Relevant works published in 16 scientific journals were analyzed globally and by continent with respect of topics, species, and families. -

Temporal Changes in the Diet of Two Sympatric Carnivorous Mammals in a Protected Area of South–Central Chile Affected by a Mixed–Severity Forest Fire
Animal Biodiversity and Conservation 43.2 (2020) 177 Temporal changes in the diet of two sympatric carnivorous mammals in a protected area of south–central Chile affected by a mixed–severity forest fire A. H. Zúñiga, J. R. Rau, V. Fuenzalida, A. Fuentes–Ramírez Zúñiga, A. H., Rau, J. R., Fuenzalida, V., Fuentes–Ramírez, A., 2020. Temporal changes in the diet of two sympatric carnivorous mammals in a protected area of south–central Chile affected by a mixed–severity forest fire. Animal Biodiversity and Conservation, 43.2: 177–186, Doi: https://doi.org/10.32800/abc.2020.43.0177 Abstract Temporal changes in the diet of two sympatric carnivorous mammals in a protected area of south–central Chile affected by a mixed–severity forest fire. Fire is a significant disruptive agent in various ecosystems around the world. It can affect the availability of resources in a given area, modulating the interaction between competing species. We studied the diet of the culpeo fox (Lycalopex culpaeus) and cougar (Puma concolor) for two consecutive years in a protected area of southern–central Chile which was affected by a wildfire. Significant differences were observed in the dietary pattern between the two species, showing their trophic segregation. In the two years of the study, the predominant prey for cougar was an exotic species, the European hare (Lepus europaeus), implying a simplification of its trophic spectrum with respect to that reported in other latitudes. The ecological consequences related to this scenario are discussed. Key words: Dietary overlap, Predation, Post–fire dynamics, Microhabitat, Rodent cycles, Selectivity Resumen Cambios temporales en la dieta de dos mamíferos carnívoros simpátridas en una zona protegida del centro y el sur de Chile afectada por un incendio forestal de intensidad desigual. -

University of Florida Thesis Or Dissertation Formatting
UNDERSTANDING CARNIVORAN ECOMORPHOLOGY THROUGH DEEP TIME, WITH A CASE STUDY DURING THE CAT-GAP OF FLORIDA By SHARON ELIZABETH HOLTE A DISSERTATION PRESENTED TO THE GRADUATE SCHOOL OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY UNIVERSITY OF FLORIDA 2018 © 2018 Sharon Elizabeth Holte To Dr. Larry, thank you ACKNOWLEDGMENTS I would like to thank my family for encouraging me to pursue my interests. They have always believed in me and never doubted that I would reach my goals. I am eternally grateful to my mentors, Dr. Jim Mead and the late Dr. Larry Agenbroad, who have shaped me as a paleontologist and have provided me to the strength and knowledge to continue to grow as a scientist. I would like to thank my colleagues from the Florida Museum of Natural History who provided insight and open discussion on my research. In particular, I would like to thank Dr. Aldo Rincon for his help in researching procyonids. I am so grateful to Dr. Anne-Claire Fabre; without her understanding of R and knowledge of 3D morphometrics this project would have been an immense struggle. I would also to thank Rachel Short for the late-night work sessions and discussions. I am extremely grateful to my advisor Dr. David Steadman for his comments, feedback, and guidance through my time here at the University of Florida. I also thank my committee, Dr. Bruce MacFadden, Dr. Jon Bloch, Dr. Elizabeth Screaton, for their feedback and encouragement. I am grateful to the geosciences department at East Tennessee State University, the American Museum of Natural History, and the Museum of Comparative Zoology at Harvard for the loans of specimens. -

Evolutionary History of Carnivora (Mammalia, Laurasiatheria) Inferred
bioRxiv preprint doi: https://doi.org/10.1101/2020.10.05.326090; this version posted October 5, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. This article is a US Government work. It is not subject to copyright under 17 USC 105 and is also made available for use under a CC0 license. 1 Manuscript for review in PLOS One 2 3 Evolutionary history of Carnivora (Mammalia, Laurasiatheria) inferred 4 from mitochondrial genomes 5 6 Alexandre Hassanin1*, Géraldine Véron1, Anne Ropiquet2, Bettine Jansen van Vuuren3, 7 Alexis Lécu4, Steven M. Goodman5, Jibran Haider1,6,7, Trung Thanh Nguyen1 8 9 1 Institut de Systématique, Évolution, Biodiversité (ISYEB), Sorbonne Université, 10 MNHN, CNRS, EPHE, UA, Paris. 11 12 2 Department of Natural Sciences, Faculty of Science and Technology, Middlesex University, 13 United Kingdom. 14 15 3 Centre for Ecological Genomics and Wildlife Conservation, Department of Zoology, 16 University of Johannesburg, South Africa. 17 18 4 Parc zoologique de Paris, Muséum national d’Histoire naturelle, Paris. 19 20 5 Field Museum of Natural History, Chicago, IL, USA. 21 22 6 Department of Wildlife Management, Pir Mehr Ali Shah, Arid Agriculture University 23 Rawalpindi, Pakistan. 24 25 7 Forest Parks & Wildlife Department Gilgit-Baltistan, Pakistan. 26 27 28 * Corresponding author. E-mail address: [email protected] bioRxiv preprint doi: https://doi.org/10.1101/2020.10.05.326090; this version posted October 5, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. This article is a US Government work. -

Activity Patterns in Sympatric Carnivores in the Nahuelbuta Mountain Range, Southern-Central Chile
Mammalia 2016; aop Alfredo H. Zúñiga*, Jaime E. Jiménez and Pablo Ramírez de Arellano Activity patterns in sympatric carnivores in the Nahuelbuta Mountain Range, southern-central Chile DOI 10.1515/mammalia-2015-0090 Received June 24, 2015; accepted August 26, 2016 Introduction Abstract: Species interactions determine the structure of Species interactions are one of the most studied topics in biological communities. In particular, interference behav- community ecology, as interspecific behavior can largely ior is critical as dominant species can displace subordi- determine the composition and structure of community nate species depending on local ecological conditions. In assemblages (Case and Gilpin 1974). For carnivores, inter- carnivores, the outcome of interference may have impor- specific interactions are particularly relevant because of tant consequences from the point of view of conservation, their role in top-down control in terrestrial ecosystems especially when vulnerable species are the ones suffering (Terborgh and Winter 1980). Nevertheless, given the key displacement. Using 24 baited camera traps and a sam- role of consumers and through trophic cascades, changes pling effort of 2821 trap nights, we examined the activ- in the environment could promote an increase of medium- ity patterns and spatial overlap of an assemblage of five sized carnivores or mesopredators, due to top predator sympatric carnivores in the Nahuelbuta Mountain Range, removal (Prange and Gehrt 2007) which can cause sub- in southern-central Chile. In this forested landscape we stantial changes in the dynamics of interaction among found predominantly nocturnal activity in all species, but sympatric species (Kamler et al. 2013), with adverse not for the puma (Puma concolor) and to a lesser extent, effects on subordinate species. -

The Ecological Effects of Providing Resource Subsidies to Predators
Global Ecology and Biogeography, (Global Ecol. Biogeogr.) (2014) bs_bs_banner RESEARCH The ecological effects of providing REVIEW resource subsidies to predators Thomas M. Newsome1,2*, Justin A. Dellinger3, Chris R. Pavey4, William J. Ripple2, Carolyn R. Shores3, Aaron J. Wirsing3 and Christopher R. Dickman1 1Desert Ecology Research Group, School of ABSTRACT Biological Sciences, University of Sydney, Aim Predators often have important roles in structuring ecosystems via their Sydney, NSW 2006, Australia, 2Trophic Cascades Program, Department of Forest effects on each other and on prey populations. However, these effects may be altered Ecosystems and Society, Oregon State in the presence of anthropogenic food resources, fuelling debate about whether the University, Corvallis, OR 97331, USA, 3School availability of such resources could alter the ecological role of predators. Here, we of Environmental and Forest Sciences, review the extent to which human-provided foods are utilised by terrestrial mam- University of Washington, Seattle, WA 98195, malian predators (> 1 kg) across the globe. We also assess whether these resources USA, 4CSIRO Land and Water Flagship, PO have a direct impact on the ecology and behaviour of predators and an indirect Box 2111, Alice Springs, NT 0871, Australia impact on other co-occurring species. Location Global. Methods Data were derived from searches of the published literature. To sum- marise the data we grouped studies based on the direct and indirect effects of resource subsidies on predators and co-occurring species. We then compared the types of predators accessing these resources by grouping species taxonomically and into the following categories: (1) domesticated species, (2) mesopredators and (3) top predators. -

Ontogenetic Origins of Cranial Convergence Between the Extinct Marsupial Thylacine and Placental Gray Wolf ✉ ✉ Axel H
ARTICLE https://doi.org/10.1038/s42003-020-01569-x OPEN Ontogenetic origins of cranial convergence between the extinct marsupial thylacine and placental gray wolf ✉ ✉ Axel H. Newton 1,2,3 , Vera Weisbecker 4, Andrew J. Pask2,3,5 & Christy A. Hipsley 2,3,5 1234567890():,; Phenotypic convergence, describing the independent evolution of similar characteristics, offers unique insights into how natural selection influences developmental and molecular processes to generate shared adaptations. The extinct marsupial thylacine and placental gray wolf represent one of the most extraordinary cases of convergent evolution in mammals, sharing striking cranial similarities despite 160 million years of independent evolution. We digitally reconstructed their cranial ontogeny from birth to adulthood to examine how and when convergence arises through patterns of allometry, mosaicism, modularity, and inte- gration. We find the thylacine and wolf crania develop along nearly parallel growth trajec- tories, despite lineage-specific constraints and heterochrony in timing of ossification. These constraints were found to enforce distinct cranial modularity and integration patterns during development, which were unable to explain their adult convergence. Instead, we identify a developmental origin for their convergent cranial morphologies through patterns of mosaic evolution, occurring within bone groups sharing conserved embryonic tissue origins. Inter- estingly, these patterns are accompanied by homoplasy in gene regulatory networks asso- ciated with neural crest cells, critical for skull patterning. Together, our findings establish empirical links between adaptive phenotypic and genotypic convergence and provides a digital resource for further investigations into the developmental basis of mammalian evolution. 1 School of Biomedical Sciences, Monash University, Melbourne, VIC, Australia. 2 School of BioSciences, The University of Melbourne, Melbourne, VIC, Australia.