Spider Genus Hispo (Araneae : Salticidae)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Open PDF Printout of This File
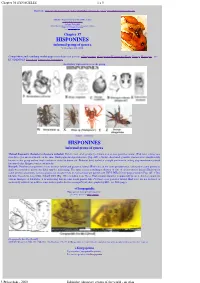
Chapter 36 SYNAGELES 1 z 5 Main links: Title page Introduction & guides INDEXES_chapters_&_genera Open PDF printout of this file Salticidae (Araneae) genera of the world - an atlas (unfinished manuscript) by Jerzy Prószy ński Professor Emeritus, Museum and Institute of Zoology, Polish Academy of Sciences, ul. Wilcza 63, 00-679 Warsaw, POLAND e-mail: [email protected] Chapter 37 HISPONINES informal group of genera Version June 3rd, 2020. symbol of the supragroup HISPONINES Composition and searching on this page (including fossil genera): +Gorgopsidis : +Gorgopsina Hermotimus Hispo Jerzego Massagris - see EUODENINES Tomobella Tomocyrba Tomomingi Exemplary representatives of the group Source - see below HISPONINES informal group of genera Mutual diagnostic characters of genera included . Differs from other groups by minute size of eyes posterior lateral (IInd row), sitting very closely to eyes anterior lateral, on the same black pigmented protuberance (Fig. 49J1). On the other hand, genitalic characters are insufficiently known in this group and no final conclusion could be drawn yet. Embolus bent, coiled or straight and minute, sitting atop membranous distal haematodocha. Epigyne varies extensively. Remark. Peculiar arrangement of eyes anterior lateral and posterior lateral (IInd row) on the same protuberance, followed in some genera by shallow constriction across eye field, may be misleading. The same concerns striking reduction of size of eyes posterior lateral (IInd row) in some genera classified to various groups, for instance eyes in Lystrocteissa are paired with HYLLINES[?] like palp structure (Figs 14L, 17G), likewise Tomobella fotsy Szüts, Scharff 2009 (Fig. 49I) resembles some Neon . That somatic character is apparently prone to develop parallel in various lineages of Salticidae, it is interesting that in some fossil genera (like Prolinus ) eyes posterior lateral (IInd row) are not reduced, or moderately reduced (as in Hisponinae amber spider shown on magnificent photographs by Hill - see Title page). -

Jerzego, a New Hisponine Jumping Spider from Borneo (Araneae: Salticidae)
Zootaxa 3852 (5): 569–578 ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ Article ZOOTAXA Copyright © 2014 Magnolia Press ISSN 1175-5334 (online edition) http://dx.doi.org/10.11646/zootaxa.3852.5.5 http://zoobank.org/urn:lsid:zoobank.org:pub:85C58188-05CE-4EED-A371-8AFE0E8148C8 Jerzego, a new hisponine jumping spider from Borneo (Araneae: Salticidae) WAYNE P. MADDISON1,3 & EDYTA K. PIASCIK2 1Departments of Zoology and Botany and Beaty Biodiversity Museum, University of British Columbia, 6270 University Boulevard, Vancouver, British Columbia, V6T 1Z4, Canada. E-mail: [email protected] 2Department of Zoology, University of British Columbia, 6270 University Boulevard, Vancouver, British Columbia, V6T 1Z4, Canada. E-mail: [email protected] 3Corresponding author Abstract A new genus and species of hisponine jumping spider from Sarawak, Jerzego corticicola Maddison sp. nov. are described, representing one of the few hisponine jumping spiders known from Asia, and the only whose male is known. Although similar to the primarily-Madagascan genus Hispo in having an elongate and flat body, sequences of 28s and 16sND1 genes indicate that Jerzego is most closely related to Massagris and Tomomingi, a result consistent with morphology. Females of Jerzego and other genera of Hisponinae were found to have an unusual double copulatory duct, which appears to be a synapomorphy of the subfamily. Two species are transferred from Hispo, Jerzego bipartitus (Simon) comb. nov. and Jer- zego alboguttatus (Simon) comb. nov. Diagnostic illustrations -

SA Spider Checklist
REVIEW ZOOS' PRINT JOURNAL 22(2): 2551-2597 CHECKLIST OF SPIDERS (ARACHNIDA: ARANEAE) OF SOUTH ASIA INCLUDING THE 2006 UPDATE OF INDIAN SPIDER CHECKLIST Manju Siliwal 1 and Sanjay Molur 2,3 1,2 Wildlife Information & Liaison Development (WILD) Society, 3 Zoo Outreach Organisation (ZOO) 29-1, Bharathi Colony, Peelamedu, Coimbatore, Tamil Nadu 641004, India Email: 1 [email protected]; 3 [email protected] ABSTRACT Thesaurus, (Vol. 1) in 1734 (Smith, 2001). Most of the spiders After one year since publication of the Indian Checklist, this is described during the British period from South Asia were by an attempt to provide a comprehensive checklist of spiders of foreigners based on the specimens deposited in different South Asia with eight countries - Afghanistan, Bangladesh, Bhutan, India, Maldives, Nepal, Pakistan and Sri Lanka. The European Museums. Indian checklist is also updated for 2006. The South Asian While the Indian checklist (Siliwal et al., 2005) is more spider list is also compiled following The World Spider Catalog accurate, the South Asian spider checklist is not critically by Platnick and other peer-reviewed publications since the last scrutinized due to lack of complete literature, but it gives an update. In total, 2299 species of spiders in 67 families have overview of species found in various South Asian countries, been reported from South Asia. There are 39 species included in this regions checklist that are not listed in the World Catalog gives the endemism of species and forms a basis for careful of Spiders. Taxonomic verification is recommended for 51 species. and participatory work by arachnologists in the region. -
The Deep Phylogeny of Jumping Spiders (Araneae, Salticidae)
A peer-reviewed open-access journal ZooKeys 440: 57–87 (2014)The deep phylogeny of jumping spiders( Araneae, Salticidae) 57 doi: 10.3897/zookeys.440.7891 RESEARCH ARTICLE www.zookeys.org Launched to accelerate biodiversity research The deep phylogeny of jumping spiders (Araneae, Salticidae) Wayne P. Maddison1,2, Daiqin Li3,4, Melissa Bodner2, Junxia Zhang2, Xin Xu3, Qingqing Liu3, Fengxiang Liu3 1 Beaty Biodiversity Museum and Department of Botany, University of British Columbia, Vancouver, British Columbia, V6T 1Z4 Canada 2 Department of Zoology, University of British Columbia, Vancouver, British Columbia, V6T 1Z4 Canada 3 Centre for Behavioural Ecology & Evolution, College of Life Sciences, Hubei University, Wuhan 430062, Hubei, China 4 Department of Biological Sciences, National University of Singa- pore, 14 Science Drive 4, Singapore 117543 Corresponding author: Wayne P. Maddison ([email protected]) Academic editor: Jeremy Miller | Received 13 May 2014 | Accepted 6 July 2014 | Published 15 September 2014 http://zoobank.org/AFDC1D46-D9DD-4513-A074-1C9F1A3FC625 Citation: Maddison WP, Li D, Bodner M, Zhang J, Xu X, Liu Q, Liu F (2014) The deep phylogeny of jumping spiders (Araneae, Salticidae). ZooKeys 440: 57–87. doi: 10.3897/zookeys.440.7891 Abstract In order to resolve better the deep relationships among salticid spiders, we compiled and analyzed a mo- lecular dataset of 169 salticid taxa (and 7 outgroups) and 8 gene regions. This dataset adds many new taxa to previous analyses, especially among the non-salticoid salticids, as well as two new genes – wingless and myosin heavy chain. Both of these genes, and especially the better sampled wingless, confirm many of the relationships indicated by other genes. -

Arachnida, Araneae) of the Savanna Biome in South Africa
The faunistic diversity of spiders (Arachnida, Araneae) of the Savanna Biome in South Africa S.H. Foord1, A.S. Dippenaar-Schoeman2& C.R Haddad3 1Department of Zoology, Centre for Invasion Biology, University of Venda, Private Bag X5050, Thohoyandou, 0950, South Africa 2 ARC-Plant Protection Research Institute, Private Bag X134, Queenswood 0121/Department of Zoology &Entomology, University of Pretoria, Pretoria 0001, South Africa 3Department of Zoology & Entomology, University of the Free State, P.O. Box 339, Bloemfontein 9300, South Africa Invertebrates include more than 80% of all animals, yet they are severely under-represented in studies of southern African diversity. Site biodiversity estimates that do not consider invertebrates, not only omit the greatest part of what they are attempting to measure, but also ignore major contributors to essential ecosystem processes. All available information on spider species distribution in the South African Savanna Biome was compiled. This is the largest biome in the country,occupying over one third of the surface area.A total of 1230 species represented by 381genera and 62 families are known from the biome. The last decade has seen an exponential growth in the knowledge of the group in South Africa, but there certainly are several more species that have to be discovered, and the distribution patterns of those listed are partly unknown. Information is summarized for all quarter degree squares of the biome and reveals considerable inequalities in knowledge. At a large scale the eastern region is much better surveyed than the western parts, but at smaller scales throughout the region, several areas have little information. -

Archiv Für Naturgeschichte
ZOBODAT - www.zobodat.at Zoologisch-Botanische Datenbank/Zoological-Botanical Database Digitale Literatur/Digital Literature Zeitschrift/Journal: Archiv für Naturgeschichte Jahr/Year: 1899 Band/Volume: 65-2_2 Autor(en)/Author(s): Lucas Robert Artikel/Article: Arachnida. 939-1000 © Biodiversity Heritage Library, http://www.biodiversitylibrary.org/; www.zobodat.at Arachnida. Bearbeitet von Dr. Robert Lucas in Reinickendorf-Schönholz. 1. Scorpiones. A. Allgemeines (Autoren, alphabetisch). Barthe, E. 1897. Scorpio occitanicus. Miscell. Entom. Vol. 5 p.ll8. Berlese. Acari, Myriopoda et Scorpiones hucusque in Italia re- perta. Parts LXXXV—XCTI, Padua, 8», 1898. Birula, A. (Titel p. 1089 sub No. 2 d. vor. Ber.) behandelt 3 (1 n. sp.); n.: Buthus fusciis; n. subsp. : Buthus caucasicus Prze- walskii. Brindley, H. H. (Titel Hft. I dieses Ber. p. 928). Cambridge, 0. P. Titel siehe p. 945 sub No. 2 dieses Berichts. Kowalevsky, AI. (Titel p. 1089 des vor. Ber.) wird referirt von R. Monti, Boll. Scientif. (Maggi, Zoja) Ann. 19 No. 4 p. 126—8. Kraepelin, Karl (1). Ueber die Linne'schen Arten der Gattung Scorpio. Eine Nomenklaturstudie. Zoolog. Anz. 21. Bd. p. 438—441, 444-447. Von den Linne'schen Sc.-Ty[)en nur 4 Exempl, vorhanden: Sc. afer, americanus, afer /? u. europaeus. cf. System. Theil. — (2). Entgegnung betreff, die Linne'schen Arten der Gattung Scorpio. t. c. p. 648^—650. — Bezieht sich auf Lönnberg. Lönnberg, Einar (1). A Revision of the Linnean Type specimens of Scorpions and Pedipalps in the Zoological Museum of the Royal University at Upsala. Ann. Nat. Hist. (7) vol. 1 p. 82—9. — (2). Noch einmal über die Linne'schen Arten der Gattung Scorpio. -

The Evolution of Jumping Spiders (Araneae: Salticidae): a Review
Peckhamia 75.1 Evolution of jumping spiders 1 PECKHAMIA 75.1, 17 August 2009, 1―7 ISSN 1944―8120 The evolution of jumping spiders (Araneae: Salticidae): a review David E. Hill1 and David B. Richman2 1 213 Wild Horse Creek Drive, Simpsonville, South Carolina 29680 2 Corresponding author: [email protected] Department of Entomology, Plant Pathology and Weed Science, New Mexico State University, Las Cruces, NM 88003 The following article was written as part of the yearlong celebration of the 200th birthday of Charles Darwin and the 150th Anniversary of the publication of his Origin of Species. Abstract The evolution of the family Salticidae (jumping spiders) has always puzzled arachnologists, but with modern DNA and morphological analysis, using cladistic techniques, a pattern is starting to emerge. The Salticidae apparently originated in the late Cretaceous and were derived from one of the other RTA dionychan clades, which include Philodromidae, Thomisidae, Miturgidae, Anyphaenidae, Gnaphosidae and related groups. Speciation appears to have been well along by the Oligocene―Miocene and may have been spurred on by the warm conditions of the late Paleocene―Eocene. At this point it seems wise to gather the current evidence and thinking on the origin and evolution of jumping spiders and the more “typical” Salticoida together and to give an overview of where the research currently stands. This is the main purpose of the current review. Introduction With more than 5,000 described species (Platnick 2009), the jumping spiders (Salticidae) arguably represent the most diverse family of modern spiders. Although we do not have any scientific standards for comparison of evolutionary diversity, those familiar with salticids might consider the group to be at least as diverse as the birds (Class Aves), and perhaps just as ancient in their origin. -

The Arthropod Fauna of Oak (Quercus Spp., Fagaceae) Canopies in Norway
diversity Article The Arthropod Fauna of Oak (Quercus spp., Fagaceae) Canopies in Norway Karl H. Thunes 1,*, Geir E. E. Søli 2, Csaba Thuróczy 3, Arne Fjellberg 4, Stefan Olberg 5, Steffen Roth 6, Carl-C. Coulianos 7, R. Henry L. Disney 8, Josef Starý 9, G. (Bert) Vierbergen 10, Terje Jonassen 11, Johannes Anonby 12, Arne Köhler 13, Frank Menzel 13 , Ryszard Szadziewski 14, Elisabeth Stur 15 , Wolfgang Adaschkiewitz 16, Kjell M. Olsen 5, Torstein Kvamme 1, Anders Endrestøl 17, Sigitas Podenas 18, Sverre Kobro 1, Lars O. Hansen 2, Gunnar M. Kvifte 19, Jean-Paul Haenni 20 and Louis Boumans 2 1 Norwegian Institute of Bioeconomy Research (NIBIO), Department Invertebrate Pests and Weeds in Forestry, Agriculture and Horticulture, P.O. Box 115, NO-1431 Ås, Norway; [email protected] (T.K.); [email protected] (S.K.) 2 Natural History Museum, University of Oslo, P.O. Box 1172 Blindern, NO-0318 Oslo, Norway; [email protected] (G.E.E.S.); [email protected] (L.O.H.); [email protected] (L.B.) 3 Malomarok, u. 27, HU-9730 Köszeg, Hungary; [email protected] 4 Mågerøveien 168, NO-3145 Tjøme, Norway; [email protected] 5 Biofokus, Gaustadalléen 21, NO-0349 Oslo, Norway; [email protected] (S.O.); [email protected] (K.M.O.) 6 University Museum of Bergen, P.O. Box 7800, NO-5020 Bergen, Norway; [email protected] 7 Kummelnäsvägen 90, SE-132 37 Saltsjö-Boo, Sweden; [email protected] 8 Department of Zoology, University of Cambridge, Downing St., Cambridge CB2 3EJ, UK; [email protected] 9 Institute of Soil Biology, Academy of Sciences of the Czech Republic, Na Sádkách 7, CZ-37005 Ceskˇ é Budˇejovice,Czech Republic; [email protected] Citation: Thunes, K.H.; Søli, G.E.E.; 10 Netherlands Food and Consumer Product Authority, P.O. -

Testing a Rapid Sampling Protocol
Page 1 of 15 Original Research Diversity of non-acarine arachnids of the Ophathe Game Reserve, South Africa: Testing a rapid sampling protocol Authors: As part of the second phase of the South African National Survey of Arachnida (SANSA), 1 Charles R. Haddad field surveys were conducted in many degree-square grids throughout the country using a Anna S. Dippenaar- Schoeman2,3 standardised rapid sampling protocol. This study reports on the arachnid diversity of the Ophathe Game Reserve (OGR) in northern KwaZulu-Natal, as found during a preliminary Affiliations: survey in June 2007 (mid winter) and a SANSA field survey in October 2008 (mid spring) in 1Department of Zoology & Entomology, University of four representative habitats. The SANSA survey included seven sampling methods: pitfalls, the Free State, South Africa beating, sweep-netting, litter sifting, hand collecting, night collecting and Winkler traps. A total of 282 species in six arachnid orders were collected during the two surveys, of which spiders 2 Biosystematics, Arachnology, were the most species-rich order (268 species in 47 families). The SANSA survey yielded 966 ARC – Plant Protection Research Institute, adult arachnids, representing six orders and 197 species, with a further 67 species represented South Africa only by immatures. Although adult arachnid abundance (n) differed considerably between the four habitats (range: 156–321), adult species richness (S ) was less variable (range: 65–85). 3Department of Zoology & obs Entomology, University of These survey results are comparable with several longer-term surveys in the Savanna biome, Pretoria, South Africa and indicate that the SANSA sampling protocol can yield an impressive diversity of arachnids during a relatively short period of sampling, with a high level of coverage (> 0.8 for sites Correspondence to: and most sampling methods) and moderate levels of sample completion for adults (> 0.55 Charles Haddad for all sites), despite logistical and temporal challenges. -

A New Species of the Hisponine Jumping Spiders from India (Araneae: Salticidae), with Some Observations on Its Life History
Arthropoda Selecta 28(1): 113–124 © ARTHROPODA SELECTA, 2019 A new species of the hisponine jumping spiders from India (Araneae: Salticidae), with some observations on its life history Íîâûé âèä ïàóêîâ-ñêàêóí÷èêîâ õèñïîíèí èç Èíäèè (Araneae: Salticidae), ñ íåêîòîðûìè íàáëþäåíèÿìè íàä åãî îáðàçîì æèçíè Rajesh V. Sanap1, John T.D. Caleb2,*, Anuradha Joglekar3 Ð.Â. Ñàíàï1, Äæ.Ò.Ä. Êàëåá2,*, À. Äæîãëåêàð3 1 National Center for Biological Sciences, Tata Institute of Fundamental Research, Bangaluru – 560 065, Karnataka, India 2 Zoological Survey of India, Prani Vigyan Bhavan, M-Block, New Alipore, Kolkata – 700 053, West Bengal, India. Email: [email protected] 3 706, Piccadilly 3, Royal Palms Estate, Aarey Milk Colony, Goregaon East, Mumbai – 400 065, Maharashtra, India *Corresponding author KEY WORDS: Aranei, bionomics, description, taxonomy, Mumbai, Jerzego. КЛЮЧЕВЫЕ СЛОВА: Aranei, биономика, описание, таксономия, Мумбай, Jerzego. ABSTRACT. A new species of the hisponine jump- described. In addition, some observations on the mat- ing spider genus Jerzego Maddison, 2014 — Jerzego ing behaviour, oviposition and postembryonic devel- sunillimaye sp.n. (#$) — is described. The species opment are documented. was found inhabiting the jungle floor of Aarey Milk Colony, Mumbai (Maharashtra, India). A detailed de- Material and methods scription and diagnostic illustrations of the species along with observations of mating, ovipostion, and postem- During June–July 2017, eight females and two males bryonic development are presented. were hand-collected. Of the collected females, four How to cite this article: Sanap R.V., Caleb J.T.D., were used for a taxonomic study, two for observing Joglekar A. 2019. A new species of the hisponine mating behaviour, and two for observing the produc- jumping spiders from India (Araneae: Salticidae), with tion of egg sacs and further postembryonic develop- some observations on its life history // Arthropoda ment of their offspring. -

Species Conservation Profiles of a Random Sample of World Spiders III: Oecobiidae to Salticidae
Biodiversity Data Journal 6: e27004 doi: 10.3897/BDJ.6.e27004 Species Conservation Profiles Species conservation profiles of a random sample of world spiders III: Oecobiidae to Salticidae Sini Seppälä‡,§, Sérgio Henriques|,§, Michael L Draney¶,§, Stefan Foord #,§, Alastair T Gibbons¤,§, Luz A Gomez«,§, Sarah Kariko »,§, Jagoba Malumbres-Olarte˄,˅,§, Marc Milne ¦,§,ˀ,§ Cor J Vink , Pedro Cardoso‡,§ ‡ Finnish Museum of Natural History, University of Helsinki, Helsinki, Finland § IUCN SSC Spider & Scorpion Specialist Group, Helsinki, Finland | University College London, London, United Kingdom ¶ University of Wisconsin-Green Bay, Green Bay, United States of America # University of Venda, Venda, South Africa ¤ University of Nottingham, Nottingham, United Kingdom « Universidad Nacional de Colombia, Bogotá, Colombia » Museum of Comparative Zoology, Harvard University, Cambridge, United States of America ˄ University of Copenhagen, Copenhagen, Denmark ˅ University of Barcelona, Barcelona, Spain ¦ University of Indianapolis, Indianapolis, United States of America ˀ Canterbury Museum, Christchurch, New Zealand Corresponding author: Pedro Cardoso (pedro.cardoso@helsinki.fi) Academic editor: Pavel Stoev Received: 27 May 2018 | Accepted: 01 Aug 2018 | Published: 03 Aug 2018 Citation: Seppälä S, Henriques S, Draney ML, Foord S, Gibbons AT, Gomez LA, Kariko S, Malumbres-Olarte J, Milne M, Vink CJ, Cardoso P (2018) Species conservation profiles of a random sample of world spiders III: Oecobiidae to Salticidae. Biodiversity Data Journal 6: e27004. https://doi.org/10.3897/BDJ.6.e27004 ZooBank: urn:lsid:zoobank.org:pub:40C3800F-B903-4786-9261-C99BD78DA017 Abstract Background The IUCN Red List of Threatened Species is the most widely used information source on the extinction risk of species. One of the uses of the Red List is to evaluate and monitor the state of biodiversity and a possible approach for this purpose is the Red List Index (RLI). -

Hymenovaria 17
nummer 17 november 2018 Nieuwsbrief Sectie Hymenoptera Nederlandse Entomologische Vereniging In dit nummer onder meer: Veldobservaties Rafelige of gladde gaten? Een onderzoekje naar bijenhotels Inventarisatie en monitoring van bijen, hoe doe je dat? Broedcellen van Isodontia mexicana Een bijdrage over de familie Platygastridae in Nederland nr. 17, november 2018 41 nr. 17, november 2018 ISSN 1387-1773 Foto voorpagina: Isodontia mexicana met prooi. Foto: Pieter van Breugel. Nieuwsbrief sectie Hymenoptera van de Nederlandse Entomologische Vereniging Vormgeving: Jan Smit. Redactie J. D’Haeseleer, T. Peeters, J. Smit, E. van der Spek Redactieadres Voermanstraat 14, 6921 NP Duiven e-mail: [email protected] Website www.hymenovaria.nl Redactioneel Dit nummer van onze nieuwsbrief is weer zeer goed De literatuurmand puilt uit, met HymenoBiblio 2017, gevuld en erg gevarieerd. Allereerst de aankondiging een viertal boekbesprekingen door Hans Nieuwen- van een excursie in het voorjaar. huijsen, Jan Smit en Theo Peeters, daarnaast geven Verder een zevental ‘Veldobservaties’. Regina Oors en Erik van der Spek inkijk in ‘Resultaten van onderzoek’. Bij de ‘Artikelen’ doet Menno Reemer verslag van een onderzoekje naar nestblokken met gladde en rafelige Bij de ‘Oproepen’ wordt gevraagd de bijdrage voor gaten, Peter Megens heeft twee bijzondere bijen in het komend jaar (2019) over te maken. Verder de vraag binnenland teruggevonden, Erik van der Spek zet alle om ‘Leuke waarnemingen en vangsten uit 2018’ op te methoden van bijeninventarisatie en –monitoring op sturen. Erik van der Spek wil graag de gegevens van een rij, Pieter van Breugel geeft ons een inkijk in de deelnemers aan de excursies van de sectie naar de Peel broedcellen van Isodontia mexicana , Martijn Kos heeft en het Hollands Duin ontvangen.