Vertical Gaze Palsy and Selective Unilateral Medial Longitudinal
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
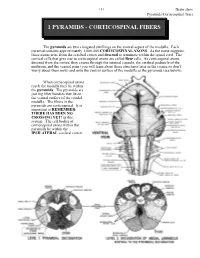
Corticospinal Fibers
151 Brain stem Pyramids/Corticospinal Tract 1 PYRAMIDS - CORTICOSPINAL FIBERS The pyramids are two elongated swellings on the ventral aspect of the medulla. Each pyramid contains approximately 1,000,000 CORTICOSPINAL AXONS. As the name suggests, these axons arise from the cerebral cortex and descend to terminate within the spinal cord. The cortical cells that give rise to corticospinal axons are called Betz cells. As corticospinal axons descend from the cortex, they course through the internal capsule, the cerebral peduncle of the midbrain, and the ventral pons (you will learn about these structures later in the course so don’t worry about them now) and onto the ventral surface of the medulla as the pyramids (see below). When corticospinal axons reach the medulla they lie within the pyramids. The pyramids are just big fiber bundles that lie on the ventral surface of the caudal medulla. The fibers in the pyramids are corticospinal. It is important to REMEMBER: THERE HAS BEEN NO CROSSING YET! in this system. The cell bodies of corticospinal axons within the pyramids lie within the IPSILATERAL cerebral cortex. Brain stem 152 Pyramids/Corticospinal Tract At the most caudal pole of the pyramids the corticospinal axons cross over the midline and now continue their descent on the contralateral (to the cell of origin) side. This crossover point is called the PYRAMIDAL DECUSSATION. The crossing fibers enter the lateral funiculus of the spinal cord where they are called the LATERAL CORTICOSPINAL TRACT (“corticospinal” is not good enough, you have to call them lateral corticospinal; LCST - remember this one??). LCST axons exit the tract to terminate upon neurons in the spinal cord gray matter along its entire length. -
L4-Physiology of Motor Tracts.Pdf
: chapter 55 page 667 Objectives (1) Describe the upper and lower motor neurons. (2) Understand the pathway of Pyramidal tracts (Corticospinal & corticobulbar tracts). (3) Understand the lateral and ventral corticospinal tracts. (4) Explain functional role of corticospinal & corticobulbar tracts. (5) Describe the Extrapyramidal tracts as Rubrospinal, Vestibulospinal, Reticulospinal and Tectspinal Tracts. The name of the tract indicate its pathway, for example Corticobulbar : Terms: - cortico: cerebral cortex. Decustation: crossing. - Bulbar: brainstem. Ipsilateral : same side. *So it starts at cerebral cortex and Contralateral: opposite side. terminate at the brainstem. CNS influence the activity of skeletal muscle through two set of neurons : 1- Upper motor neurons (UMN) 2- lower motor neuron (LMN) They are neurons of motor cortex & their axons that pass to brain stem and They are Spinal motor neurons in the spinal spinal cord to activate: cord & cranial motor neurons in the brain • cranial motor neurons (in brainstem) stem which innervate muscles directly. • spinal motor neurons (in spinal cord) - These are the only neurons that innervate - Upper motor neurons (UMN) are the skeletal muscle fibers, they function as responsible for conveying impulses for the final common pathway, the final link voluntary motor activity through between the CNS and skeletal muscles. descending motor pathways that make up by the upper motor neurons. Lower motor neurons are classified based on the type of muscle fiber the innervate: There are two UMN Systems through which 1- alpha motor neurons (UMN) control (LMN): 2- gamma motor neurons 1- Pyramidal system (corticospinal tracts ). 2- Extrapyramidal system The activity of the lower motor neuron (LMN, spinal or cranial) is influenced by: 1. -

Visualization of the Pyramidal Decussation Utilizing Diffusion
OPEN ACCESS RESEARCH Visualization of the pyramidal decussation utilizing diffusion tensor imaging: a feasibility study utilizing generalized q-sampling imaging Erik H Middlebrooks1,*, Jeffrey A Bennett1, Sharatchandra Bidari1, and Alissa Old Crow1 1Department of Radiology, University of Florida College of Medicine, Gainesville, Florida, USA *corresponding author ([email protected]) Abstract Background: The delineating point of the end of the brainstem and beginning of the spinal cord is known as the cervicomedullary junction (CMJ). This point is defined by the decussation of the pyramidal tracts. Abnormal configuration and location of the CMJ have both been implicated in disease processes such as Chiari malformation. Unfortunately, the CMJ is not directly visualized on contemporary imaging techniques. Diffusion tensor imaging (DTI) has given us the ability to directly visualize white matter tracts, but suffers from difficulties with visualizing crossing fibers. Many advanced techniques for visualizing crossing fibers utilize substantially long imaging times or non-clinical magnet strengths making clinical applicability limited. Objective: This study inves- tigates the use of generalized q-sampling imaging with diffusion decomposition on standard DTI acquisitions at 3 Tesla to demonstrate the pyramidal decussation. Methods: Three differing DTI protocols were analyzed in vivo with scan times of 17 minutes, 10 minutes, and 5.5 minutes. Data was processed with standard DTI post-processing, as well as generalized q-sampling imaging with diffusion decomposition. Results: The results of the study show that the pyramidal decussation can be reliably visualized using generalized q-sampling imaging and diffusion decomposition with scan times as low at 10 minutes. Conclusion: Utilizing generalized q-sampling post-processing, the pyramidal decussation can be reliably visualized using clinically feasible DTI sequences with scan times as low as 10 minutes. -

Auditory and Vestibular Systems Objective • to Learn the Functional
Auditory and Vestibular Systems Objective • To learn the functional organization of the auditory and vestibular systems • To understand how one can use changes in auditory function following injury to localize the site of a lesion • To begin to learn the vestibular pathways, as a prelude to studying motor pathways controlling balance in a later lab. Ch 7 Key Figs: 7-1; 7-2; 7-4; 7-5 Clinical Case #2 Hearing loss and dizziness; CC4-1 Self evaluation • Be able to identify all structures listed in key terms and describe briefly their principal functions • Use neuroanatomy on the web to test your understanding ************************************************************************************** List of media F-5 Vestibular efferent connections The first order neurons of the vestibular system are bipolar cells whose cell bodies are located in the vestibular ganglion in the internal ear (NTA Fig. 7-3). The distal processes of these cells contact the receptor hair cells located within the ampulae of the semicircular canals and the utricle and saccule. The central processes of the bipolar cells constitute the vestibular portion of the vestibulocochlear (VIIIth cranial) nerve. Most of these primary vestibular afferents enter the ipsilateral brain stem inferior to the inferior cerebellar peduncle to terminate in the vestibular nuclear complex, which is located in the medulla and caudal pons. The vestibular nuclear complex (NTA Figs, 7-2, 7-3), which lies in the floor of the fourth ventricle, contains four nuclei: 1) the superior vestibular nucleus; 2) the inferior vestibular nucleus; 3) the lateral vestibular nucleus; and 4) the medial vestibular nucleus. Vestibular nuclei give rise to secondary fibers that project to the cerebellum, certain motor cranial nerve nuclei, the reticular formation, all spinal levels, and the thalamus. -

Microstimulation of the Midbrain Tegmentum Creates Learning Signals for Saccade Adaptation
The Journal of Neuroscience, April 4, 2007 • 27(14):3759–3767 • 3759 Behavioral/Systems/Cognitive Microstimulation of the Midbrain Tegmentum Creates Learning Signals for Saccade Adaptation Yoshiko Kojima, Kaoru Yoshida, and Yoshiki Iwamoto Department of Neurophysiology, Doctoral Program in Kansei Behavioral and Brain Sciences, University of Tsukuba, Tsukuba, Ibaraki 305-8574, Japan Error signals are vital to motor learning. However, we know little about pathways that transmit error signals for learning in voluntary movements. Here we show that microstimulation of the midbrain tegmentum can induce learning in saccadic eye movements in mon- keys. Weak electrical stimuli delivered ϳ200 ms after saccades in one horizontal direction produced gradual and marked changes in saccade gain. The spatial and temporal characteristics of the produced changes were similar to those of adaptation induced by real visual error. When stimulation was applied after saccades in two different directions, endpoints of these saccades gradually shifted in the same direction in two dimensions. We conclude that microstimulation created powerful learning signals that dictate the direction of adaptive shift in movement endpoints. Our findings suggest that the error signals for saccade adaptation are conveyed in a pathway that courses through the midbrain tegmentum. Key words: motor learning; electrical stimulation; midbrain; saccade; adaptation; macaque; monkey Introduction superior colliculus, a key midbrain structure that issues saccade Motor learning ensures the accuracy of the movements we exe- signals to the premotor reticular circuitry (Scudder et al., 2002). cute daily. Vital to learning is the information about the error that The colliculus also has indirect access to the cerebellum via both results from executed movements. -

Brain Structure and Function Related to Headache
Review Cephalalgia 0(0) 1–26 ! International Headache Society 2018 Brain structure and function related Reprints and permissions: sagepub.co.uk/journalsPermissions.nav to headache: Brainstem structure and DOI: 10.1177/0333102418784698 function in headache journals.sagepub.com/home/cep Marta Vila-Pueyo1 , Jan Hoffmann2 , Marcela Romero-Reyes3 and Simon Akerman3 Abstract Objective: To review and discuss the literature relevant to the role of brainstem structure and function in headache. Background: Primary headache disorders, such as migraine and cluster headache, are considered disorders of the brain. As well as head-related pain, these headache disorders are also associated with other neurological symptoms, such as those related to sensory, homeostatic, autonomic, cognitive and affective processing that can all occur before, during or even after headache has ceased. Many imaging studies demonstrate activation in brainstem areas that appear specifically associated with headache disorders, especially migraine, which may be related to the mechanisms of many of these symptoms. This is further supported by preclinical studies, which demonstrate that modulation of specific brainstem nuclei alters sensory processing relevant to these symptoms, including headache, cranial autonomic responses and homeostatic mechanisms. Review focus: This review will specifically focus on the role of brainstem structures relevant to primary headaches, including medullary, pontine, and midbrain, and describe their functional role and how they relate to mechanisms -

(Rmtg),&Nbsp;A Gabaergic Afferent to Midbrain Dopamine Neurons
Neuron Article The Rostromedial Tegmental Nucleus (RMTg), a GABAergic Afferent to Midbrain Dopamine Neurons, Encodes Aversive Stimuli and Inhibits Motor Responses Thomas C. Jhou,1,* Howard L. Fields,2 Mark G. Baxter,3 Clifford B. Saper,4 and Peter C. Holland5 1Behavioral Neuroscience Branch, National Institute on Drug Abuse, Baltimore, MD 21224, USA 2Ernest Gallo Clinic and Research Center, University of California at San Francisco, Emeryville, CA 94608, USA 3Department of Experimental Psychology, University of Oxford, OX1 3UD Oxford, UK 4Beth Israel Deaconess Medical Center, Harvard University, Boston, MA 02215, USA 5Department of Psychological and Brain Sciences, Johns Hopkins University, Baltimore, MD 21218, USA *Correspondence: [email protected] DOI 10.1016/j.neuron.2009.02.001 SUMMARY vation strongly inhibits dopamine (DA) neurons (Christoph et al., 1986; Ji and Shepard, 2007), are activated by aversive Separate studies have implicated the lateral habe- stimuli and reward omission and inhibited by reward-predictive nula (LHb) or amygdala-related regions in processing cues or unexpected rewards (Matsumoto and Hikosaka, 2007, aversive stimuli, but their relationships to each other 2009; Geisler and Trimble, 2008). These ‘‘negative reward and to appetitive motivational systems are poorly prediction errors’’ are inverse to firing patterns in putative dopa- understood. We show that neurons in the recently minergic midbrain neurons (Mirenowicz and Schultz, 1994, 1996; identified GABAergic rostromedial tegmental Hollerman and Schultz, 1998; -

Orienting Head Movements Resulting from Electrical Microstimulation of the Brainstem Tegmentum in the Barn Owl
The Journal of Neuroscience, January 1993, 13(l): 351370 Orienting Head Movements Resulting from Electrical Microstimulation of the Brainstem Tegmentum in the Barn Owl Tom Masino and Eric I. Knudsen Department of Neurobiology, Stanford University, Stanford, California 943055401 The size and direction of orienting movements are repre- movement latency, duration, velocity, and size each dem- sented systematically as a motor map in the optic tectum of onstrated dependencies on stimulus amplitude, frequency, the barn owl (du Lac and Knudsen, 1990). The optic tectum and duration. projects to several distinct regions in the medial brainstem The data demonstrate directly that at the level of the mid- tegmentum, which in turn project to the spinal cord (Masino brain tegmentum there exists a three-dimensional Cartesian and Knudsen, 1992). This study explores the hypothesis that representation of head-orienting movements such that hor- a fundamental transformation in the neural representation izontal, vertical, and roll components of movement are en- of orienting movements takes place in the brainstem teg- coded by anatomically distinct neural circuits. The data sug- mentum. Head movements evoked by electrical microstim- gest that in the projection from the optic tectum to these ulation in the brainstem tegmentum of the alert barn owl were medial tegmental regions, the topographic code for orienting cataloged and the sites of stimulation were reconstructed movement that originates in the tectum is transformed into histologically. Movements elicited from the brainstem teg- this Cartesian code. mentum were categorized into one of six different classes: [Key words: optic tectum, superior colliculus, saccadic saccadic head rotations, head translations, facial move- head movement, brainstem tegmentum, interstitial nucleus ments, vocalizations, limb movements, and twitches. -

Why a Midbrain? Evolution, Structure and Functions Notes
The changes cause "distortions" in the basic organization of the hindbrain • Variations in relative size of parts – Huge vagal lobe of the fresh-water buffalofish [Review] – Vagal and facial lobes of the catfish [Review] – Electric fish have an enormous and specialized cerebellum. [Review] • Cell migrations from the rostral hindbrain’s alar plate— from a proliferative region called the “rhombic lip”: – To cerebellum – To pre-cerebellar cell groups – especially the cells of the pons 1 Endbrain Midbrain Cerebellum Vagal Lobe Image by MIT OpenCourseWare. Buffalofish (Carpiodes tumidus) has a specialized palatal organ for filtering the water for food; it is innervated by the vagus nerve. 2 Olfactory Stalk Primitive Endbrain Midbrain Cerebellum Facial Lobe Vagal Lobe Image by MIT OpenCourseWare. The catfish has taste receptors all over its body innervated by the facial nerve (7th cranial nerve) 3 Amiurus melas (the small catfish) Image by MIT OpenCourseWare. 4 Courtesy of MIT Press. Used with permission. Schneider, G. E. Brain structure and its Origins: In the Development and in Evolution of Behavior and the Mind. MIT Press, 2014. ISBN: 9780262026734. The enlarged cerebellum of a Mormyrid fish Fig.6-1 5 Questions, chapter 10 16) What is the meaning of the term “pons”? (See the end of chapter 5.) What is a major input, and what is the major output, of the cells of the pontine gray matter? 17) What causes quantitative distortions of the basic structural layout of the hindbrain? What is the major distortion that occurs in the development of the hindbrain of humans and other primates? 18) What is the role of the “rhombic lip” – a structure seen during the development of the rostral hindbrain? 6 The "distortions" in the basic organization of the hindbrain, continued • Variations in relative size of parts √ Huge vagal lobe of the fresh-water buffalofish √ Vagal and facial lobes of the catfish √ Electric fish have an enormous and specialized cerebellum. -

Localization and Network of Coma- Causing Brainstem Lesions: Evidence for a Human Consciousness Network
Localization and Network of Coma- Causing Brainstem Lesions: Evidence for a Human Consciousness Network The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters Citation Fischer, David B. 2016. Localization and Network of Coma-Causing Brainstem Lesions: Evidence for a Human Consciousness Network. Doctoral dissertation, Harvard Medical School. Citable link http://nrs.harvard.edu/urn-3:HUL.InstRepos:27007725 Terms of Use This article was downloaded from Harvard University’s DASH repository, and is made available under the terms and conditions applicable to Other Posted Material, as set forth at http:// nrs.harvard.edu/urn-3:HUL.InstRepos:dash.current.terms-of- use#LAA Abstract Focal brainstem lesions can disrupt arousal and cause coma, yet the exact location of the brainstem region critical to arousal and its associated network are unknown. First, we compare brainstem lesions between 12 patients with coma and 24 patients without coma to identify a region specific to coma-causing lesions. Second, we determine the network connectivity of this brainstem region and each individual coma- causing lesion using resting state functional connectivity MRI data acquired from 98 healthy subjects. Third, we evaluate the functional connectivity of this network in patients with disorders of consciousness (51 patients versus 21 controls). These analyses reveal a small, coma-specific region in the left pontine tegmentum, near the medial parabrachial nucleus. This brainstem region, and each individual coma-causing lesion, is functionally connected to the left agranular, anterior insula (AI), and pregenual anterior cingulate cortex (pACC). These cortical sites align poorly with previously defined functional networks but match the distribution of von Economo neurons (VENs). -

Quantifying Nerve Decussation Abnormalities in the Optic Chiasm T Robert J
NeuroImage: Clinical 24 (2019) 102055 Contents lists available at ScienceDirect NeuroImage: Clinical journal homepage: www.elsevier.com/locate/ynicl Quantifying nerve decussation abnormalities in the optic chiasm T Robert J. Puzniaka, Khazar Ahmadia, Jörn Kaufmannb, Andre Gouwsc, Antony B. Morlandc,d, ⁎ Franco Pestillie,1, Michael B. Hoffmanna,f,1, a Department of Ophthalmology, Otto-von-Guericke-University Magdeburg, Magdeburg, Germany b Department of Neurology, Otto-von-Guericke-University Magdeburg, Magdeburg, Germany c York Neuroimaging Centre, Department of Psychology, University of York, York, United Kingdom d York Biomedical Research Institute, University of York, York, United Kingdom e Department of Psychological and Brain Sciences, Program in Neuroscience and Program in Cognitive Science, Indiana University, Bloomington, USA f Center for Behavioral Brain Sciences, Magdeburg, Germany ARTICLE INFO ABSTRACT Keywords: Objective: The human optic chiasm comprises partially crossing optic nerve fibers. Here we used diffusion MRI Optic chiasm (dMRI) for the in-vivo identification of the abnormally high proportion of crossing fibers found intheoptic Albinism chiasm of people with albinism. Diffusion MRI Methods: In 9 individuals with albinism and 8 controls high-resolution 3T dMRI data was acquired and analyzed Functional MRI with a set of methods for signal modeling [Diffusion Tensor (DT) and Constrained Spherical Deconvolution Crossing nerves (CSD)], tractography, and streamline filtering (LiFE, COMMIT, and SIFT2). The number of crossing andnon- crossing streamlines and their weights after filtering entered ROC-analyses to compare the discriminative power of the methods based on the area under the curve (AUC). The dMRI results were cross-validated with fMRI estimates of misrouting in a subset of 6 albinotic individuals. -

Chapter 3: Internal Anatomy of the Central Nervous System
10353-03_CH03.qxd 8/30/07 1:12 PM Page 82 3 Internal Anatomy of the Central Nervous System LEARNING OBJECTIVES Nuclear structures and fiber tracts related to various functional systems exist side by side at each level of the After studying this chapter, students should be able to: nervous system. Because disease processes in the brain • Identify the shapes of corticospinal fibers at different rarely strike only one anatomic structure or pathway, there neuraxial levels is a tendency for a series of related and unrelated clinical symptoms to emerge after a brain injury. A thorough knowl- • Recognize the ventricular cavity at various neuroaxial edge of the internal brain structures, including their shape, levels size, location, and proximity, makes it easier to understand • Recognize major internal anatomic structures of the their functional significance. In addition, the proximity of spinal cord and describe their functions nuclear structures and fiber tracts explains multiple symp- toms that may develop from a single lesion site. • Recognize important internal anatomic structures of the medulla and explain their functions • Recognize important internal anatomic structures of the ANATOMIC ORIENTATION pons and describe their functions LANDMARKS • Identify important internal anatomic structures of the midbrain and discuss their functions Two distinct anatomic landmarks used for visual orientation to the internal anatomy of the brain are the shapes of the • Recognize important internal anatomic structures of the descending corticospinal fibers and the ventricular cavity forebrain (diencephalon, basal ganglia, and limbic (Fig. 3-1). Both are present throughout the brain, although structures) and describe their functions their shape and size vary as one progresses caudally from the • Follow the continuation of major anatomic structures rostral forebrain (telencephalon) to the caudal brainstem.