Decussation Geometries in the Goldfish Nervous System
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
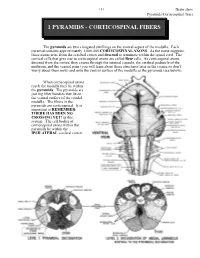
Corticospinal Fibers
151 Brain stem Pyramids/Corticospinal Tract 1 PYRAMIDS - CORTICOSPINAL FIBERS The pyramids are two elongated swellings on the ventral aspect of the medulla. Each pyramid contains approximately 1,000,000 CORTICOSPINAL AXONS. As the name suggests, these axons arise from the cerebral cortex and descend to terminate within the spinal cord. The cortical cells that give rise to corticospinal axons are called Betz cells. As corticospinal axons descend from the cortex, they course through the internal capsule, the cerebral peduncle of the midbrain, and the ventral pons (you will learn about these structures later in the course so don’t worry about them now) and onto the ventral surface of the medulla as the pyramids (see below). When corticospinal axons reach the medulla they lie within the pyramids. The pyramids are just big fiber bundles that lie on the ventral surface of the caudal medulla. The fibers in the pyramids are corticospinal. It is important to REMEMBER: THERE HAS BEEN NO CROSSING YET! in this system. The cell bodies of corticospinal axons within the pyramids lie within the IPSILATERAL cerebral cortex. Brain stem 152 Pyramids/Corticospinal Tract At the most caudal pole of the pyramids the corticospinal axons cross over the midline and now continue their descent on the contralateral (to the cell of origin) side. This crossover point is called the PYRAMIDAL DECUSSATION. The crossing fibers enter the lateral funiculus of the spinal cord where they are called the LATERAL CORTICOSPINAL TRACT (“corticospinal” is not good enough, you have to call them lateral corticospinal; LCST - remember this one??). LCST axons exit the tract to terminate upon neurons in the spinal cord gray matter along its entire length. -

Quantitative Analysis of Axon Collaterals of Single Pyramidal Cells
Yang et al. BMC Neurosci (2017) 18:25 DOI 10.1186/s12868-017-0342-7 BMC Neuroscience RESEARCH ARTICLE Open Access Quantitative analysis of axon collaterals of single pyramidal cells of the anterior piriform cortex of the guinea pig Junli Yang1,2*, Gerhard Litscher1,3* , Zhongren Sun1*, Qiang Tang1, Kiyoshi Kishi2, Satoko Oda2, Masaaki Takayanagi2, Zemin Sheng1,4, Yang Liu1, Wenhai Guo1, Ting Zhang1, Lu Wang1,3, Ingrid Gaischek3, Daniela Litscher3, Irmgard Th. Lippe5 and Masaru Kuroda2 Abstract Background: The role of the piriform cortex (PC) in olfactory information processing remains largely unknown. The anterior part of the piriform cortex (APC) has been the focus of cortical-level studies of olfactory coding, and asso- ciative processes have attracted considerable attention as an important part in odor discrimination and olfactory information processing. Associational connections of pyramidal cells in the guinea pig APC were studied by direct visualization of axons stained and quantitatively analyzed by intracellular biocytin injection in vivo. Results: The observations illustrated that axon collaterals of the individual cells were widely and spatially distrib- uted within the PC, and sometimes also showed a long associational projection to the olfactory bulb (OB). The data showed that long associational axons were both rostrally and caudally directed throughout the PC, and the intrinsic associational fibers of pyramidal cells in the APC are omnidirectional connections in the PC. Within the PC, associa- tional axons typically followed rather linear trajectories and irregular bouton distributions. Quantitative data of the axon collaterals of two pyramidal cells in the APC showed that the average length of axonal collaterals was 101 mm, out of which 79 mm (78% of total length) were distributed in the PC. -

Isolated Relative Afferent Pupillary Defect Secondary to Contralateral Midbrain Compression
OBSERVATION Isolated Relative Afferent Pupillary Defect Secondary to Contralateral Midbrain Compression Cheun Ju Chen, MD; Mia Scheufele, MD; Maushmi Sheth, MD; Amir Torabi, MD; Nick Hogan, MD, PhD; Elliot M. Frohman, MD, PhD Background: Relative afferent pupillary defects are typi- accounts for the relative afferent pupillary defect con- cally related to ipsilateral lesions within the anterior vi- tralateral to the described lesion. sual pathways. Result: Magnetic resonance imaging of the brain revealed a pineal tumor compressing the right rostral midbrain. Objective: To describe a patient who had a workup for headache and was found to have an isolated left relative Conclusion: While rare, a relative afferent pupillary de- afferent pupillary defect without any other neurological fect can occasionally occur secondary to lesions in the findings. postchiasmal pathways. In these circumstances, the pu- pillary defect will be observed to be contralateral to the Design: We review the neuroanatomy of the pupil- side of the lesion. lary light reflex pathway and emphasize the nasotem- poral bias of decussating fiber projections, which Arch Neurol. 2004;61:1451-1453 RELATIVE AFFERENT PUPIL- though retinal fibers concerned with this lary defect (RAPD) is char- reflex transmit information to both the ip- acterized by pupillary dila- silateral and contralateral midbrain, there tion upon illuminating the is a slight crossing bias, with about 53% of eye during the swinging the fibers crossing in the optic chiasm Aflashlight test. The presence of this sign sig- (chiefly derived from the nasal retina) and nifies an abnormality in the transmission 47% remaining ipsilateral. This anatomi- of light information within the pupillary cal organization of the pupillary constric- light constrictor pathway from the retina tor pathway results in the possibility of pro- to the rostral midbrain circuitry involved ducing an RAPD during illumination of the in this reflex. -

Measurement of the Normal Optic Chiasm on Coronal MR Images
Measurement of the Normal Optic Chiasm on Coronal MR Images Andrew L. Wagner, F. Reed Murtagh, Ken S. Hazlett, and John A. Arrington PURPOSE: To develop an objective method for measuring the optic chiasm and to document its normal range in size. METHODS: Measurements of the height and area of the optic chiasm, made on coronal T1-weighted MR images with the use of commercially available region-of-interest software, were obtained in 114 healthy subjects who had a total of 123 MR studies. A normal range and standard deviation were calculated, and the information was broken down by age and sex. RESULTS: The mean area of the optic chiasm was 43.7 mm2, with a standard deviation of 5.21. The mean width was 14.0 mm, with a standard deviation of 1.68. CONCLUSION: The area and width of the optic chiasm can be measured with the use of commercially available software, which allows an objective estimate of the chiasm’s size. Knowledge of the normal size range of the optic chiasm can be helpful in the early detection of some disorders. Index terms: Optic chiasm; Brain, anatomy; Brain, measurement AJNR Am J Neuroradiol 18:723–726, April 1997 The optic chiasm is an important land- months and that had been interpreted as normal. No pa- mark when interpreting magnetic resonance (MR) tient had suspected visual or endocrine abnormalities. All examinations of the brain. A small chiasm can be the examinations had been performed with a 1.5-T Gen- an indication of several disorders, the most com- eral Electric (Milwaukee, Wis) Signa or 1.5-T Siemens mon of which is septooptic dysplasia (1), and a (Cary, NC) Somatom MR system using routine imaging large chiasm can be the result of glioma, menin- protocols, with additional 3-mm T1-weighted contiguous coronal sections used for measurements. -

Visualization of the Pyramidal Decussation Utilizing Diffusion
OPEN ACCESS RESEARCH Visualization of the pyramidal decussation utilizing diffusion tensor imaging: a feasibility study utilizing generalized q-sampling imaging Erik H Middlebrooks1,*, Jeffrey A Bennett1, Sharatchandra Bidari1, and Alissa Old Crow1 1Department of Radiology, University of Florida College of Medicine, Gainesville, Florida, USA *corresponding author ([email protected]) Abstract Background: The delineating point of the end of the brainstem and beginning of the spinal cord is known as the cervicomedullary junction (CMJ). This point is defined by the decussation of the pyramidal tracts. Abnormal configuration and location of the CMJ have both been implicated in disease processes such as Chiari malformation. Unfortunately, the CMJ is not directly visualized on contemporary imaging techniques. Diffusion tensor imaging (DTI) has given us the ability to directly visualize white matter tracts, but suffers from difficulties with visualizing crossing fibers. Many advanced techniques for visualizing crossing fibers utilize substantially long imaging times or non-clinical magnet strengths making clinical applicability limited. Objective: This study inves- tigates the use of generalized q-sampling imaging with diffusion decomposition on standard DTI acquisitions at 3 Tesla to demonstrate the pyramidal decussation. Methods: Three differing DTI protocols were analyzed in vivo with scan times of 17 minutes, 10 minutes, and 5.5 minutes. Data was processed with standard DTI post-processing, as well as generalized q-sampling imaging with diffusion decomposition. Results: The results of the study show that the pyramidal decussation can be reliably visualized using generalized q-sampling imaging and diffusion decomposition with scan times as low at 10 minutes. Conclusion: Utilizing generalized q-sampling post-processing, the pyramidal decussation can be reliably visualized using clinically feasible DTI sequences with scan times as low as 10 minutes. -

Auditory and Vestibular Systems Objective • to Learn the Functional
Auditory and Vestibular Systems Objective • To learn the functional organization of the auditory and vestibular systems • To understand how one can use changes in auditory function following injury to localize the site of a lesion • To begin to learn the vestibular pathways, as a prelude to studying motor pathways controlling balance in a later lab. Ch 7 Key Figs: 7-1; 7-2; 7-4; 7-5 Clinical Case #2 Hearing loss and dizziness; CC4-1 Self evaluation • Be able to identify all structures listed in key terms and describe briefly their principal functions • Use neuroanatomy on the web to test your understanding ************************************************************************************** List of media F-5 Vestibular efferent connections The first order neurons of the vestibular system are bipolar cells whose cell bodies are located in the vestibular ganglion in the internal ear (NTA Fig. 7-3). The distal processes of these cells contact the receptor hair cells located within the ampulae of the semicircular canals and the utricle and saccule. The central processes of the bipolar cells constitute the vestibular portion of the vestibulocochlear (VIIIth cranial) nerve. Most of these primary vestibular afferents enter the ipsilateral brain stem inferior to the inferior cerebellar peduncle to terminate in the vestibular nuclear complex, which is located in the medulla and caudal pons. The vestibular nuclear complex (NTA Figs, 7-2, 7-3), which lies in the floor of the fourth ventricle, contains four nuclei: 1) the superior vestibular nucleus; 2) the inferior vestibular nucleus; 3) the lateral vestibular nucleus; and 4) the medial vestibular nucleus. Vestibular nuclei give rise to secondary fibers that project to the cerebellum, certain motor cranial nerve nuclei, the reticular formation, all spinal levels, and the thalamus. -

1. Lateral View of Lobes in Left Hemisphere TOPOGRAPHY
TOPOGRAPHY T1 Division of Cerebral Cortex into Lobes 1. Lateral View of Lobes in Left Hemisphere 2. Medial View of Lobes in Right Hemisphere PARIETAL PARIETAL LIMBIC FRONTAL FRONTAL INSULAR: buried OCCIPITAL OCCIPITAL in lateral fissure TEMPORAL TEMPORAL 3. Dorsal View of Lobes 4. Ventral View of Lobes PARIETAL TEMPORAL LIMBIC FRONTAL OCCIPITAL FRONTAL OCCIPITAL Comment: The cerebral lobes are arbitrary divisions of the cerebrum, taking their names, for the most part, from overlying bones. They are not functional subdivisions of the brain, but serve as a reference for locating specific functions within them. The anterior (rostral) end of the frontal lobe is referred to as the frontal pole. Similarly, the anterior end of the temporal lobe is the temporal pole, and the posterior end of the occipital lobe the occipital pole. TOPOGRAPHY T2 central sulcus central sulcus parietal frontal occipital lateral temporal lateral sulcus sulcus SUMMARY CARTOON: LOBES SUMMARY CARTOON: GYRI Lateral View of Left Hemisphere central sulcus postcentral superior parietal superior precentral gyrus gyrus lobule frontal intraparietal sulcus gyrus inferior parietal lobule: supramarginal and angular gyri middle frontal parieto-occipital sulcus gyrus incision for close-up below OP T preoccipital O notch inferior frontal cerebellum gyrus: O-orbital lateral T-triangular sulcus superior, middle and inferior temporal gyri OP-opercular Lateral View of Insula central sulcus cut surface corresponding to incision in above figure insula superior temporal gyrus Comment: Insula (insular gyri) exposed by removal of overlying opercula (“lids” of frontal and parietal cortex). TOPOGRAPHY T3 Language sites and arcuate fasciculus. MRI reconstruction from a volunteer. central sulcus supramarginal site (posterior Wernicke’s) Language sites (squares) approximated from electrical stimulation sites in patients undergoing operations for epilepsy or tumor removal (Ojeman and Berger). -

Cranial Nerves II, III, IV & VI (Optic, Oculomotor, Trochlear, & Abducens)
Cranial Nerves II, III, IV & VI (Optic, Oculomotor, Trochlear, & Abducens) Lecture (13) ▪ Important ▪ Doctors Notes Please check our Editing File ▪ Notes/Extra explanation ه هذا العمل مب ين بشكل أسا يس عىل عمل دفعة 436 مع المراجعة { َوَم نْ يَ َت َو َ ّكْ عَ َلْ ا َّْلل فَهُ َوْ َحْ سْ ُ ُُْ} والتدقيق وإضافة المﻻحظات وﻻ يغ ين عن المصدر اﻷسا يس للمذاكرة ▪ Objectives At the end of the lecture, students should be able to: ✓ List the cranial nuclei related to occulomotor, trochlear, and abducent nerves in the brain stem. ✓ Describe the type and site of each nucleus. ✓ Describe the site of emergence and course of these 3 nerves. ✓ Describe the important relations of oculomotor, trochlear, and abducent nerves in the orbit ✓ List the orbital muscles supplied by each of these 3 nerves. ✓ Describe the effect of lesion of each of these 3 nerves. ✓ Describe the optic nerve and visual pathway. Recall the how these nerves exit from the brain stem: Optic (does not exit from brain stem) Occulomotor: ventral midbrain (medial aspect of crus cerebri) Trochlear: dorsal midbrain (caudal to inferior colliculus) Abducent: ventral Pons (junction b/w pons & pyramid) Brain (Ventral view) Brain stem (Lateral view) Extra-Ocular Muscles 7 muscles: (ترفع جفن العين) .Levator palpebrae superioris 1- Origin: from the roof of the orbit (4) Recti muscles: *Rectus: ماشي على ( Superior rectus (upward and medially 2- الصراط (Inferior rectus (downward and medially 3- المستقيم 4- Medial rectus (medial) (medial) 5- Lateral rectus (lateral) How to remember the 2 فحركته muscles not supplied by نفس اسمه -اسمها عكس وظيفتها- :Oblique muscles (2) 6- Superior oblique (downward and laterally) Oblique: CN3? Superior oblique goes -1 منحرفOrigin: from the roof of the orbit 7- Inferior oblique (upward and laterally) up (superior) and turns around (oblique) a notch يمشي Origin: from the anterior floor or pulley and its supply is عكس كﻻمه NB. -

ACNS1721 and ACNS1723 Contouring Atlas
ACNS1721 and ACNS1723 Contouring Atlas John T. Lucas Jr.1, Stephanie M. Perkins2, David R. Raleigh3, Matthew M. Ladra4, Erin S. Murphy5, Stephanie A. Terezakis4, Thomas E. Merchant1, Daphne A. Haas-Kogan6 Shannon M. MacDonald7 1St. Jude Children’s Research Hospital, 2Washington University School of Medicine in St. Louis, 3University of California San Francisco, 4Johns Hopkins University, 5Cleveland Clinic, 6Dana-Farber Cancer Institute, 7Massachusetts General Hospital 1 Radiotherapy Planning Scans • CT Simulation: – Non-contrast treatment-planning CT scan of the entire head region. – 1.25-1.5mm slice thickness is preferred. – Immobilize patient in supine position using an immobilization device such as an Aquaplast mask over the head. • MRI-CT Fusion: – Register and fuse the relevant MRI sequences to the treatment-planning CT. – Suggested imaging type for delineation of organs at risk and target volumes are detailed in the following slides. 2 OAR General Principles • Please adhere to use of standard name terminology detailed in Section 17.9. • The following imaging is suggested for delineation of organs at risk: – Cochlea, lens: CT planning scan (bone window) – Brainstem, optic n., chiasm: Isovolumetric imaging (MPRAGE or SPGR) T1 or T2 – Body: CT planning scan – Optic Globes: CT planning scan (brain or head and neck window) 3 Required Organs at Risk Description Standard Name Goal Maximum Right Optic Nerve OpticNrv_R D50% < 5400 cGy & D0.1cc < 5600 cGy Left Optic Nerve OpticNrv_L Optic Chiasm OpticChiasm Optic Nerves & Chiasm PRV D50% < 5600 cGy & D10% < 5800 cGy – PRV Brainstem Brainstem D50% < 5240 cGy, D10% < 5600 cGy & D0.1cc < 5880 cGy Spinal Cord SpinalCord D0.1cc < 5400 cGy Right Cochlea Cochlea_R D50% < 3500 cGy (single cochlea) D50% < 2000 cGy – Preferred Left Cochlea Cochlea_L (single cochlea) Body External Unspecified Tissue Optic Globes D50% < 1000 cGy & D10% < 3500 D50% < 2000 cGy & D10% < cGy 5400 cGy 4 See ACNS1721 Protocol Section 17.8 and 17.9 for structure definitions and further detail regarding constraints. -

Clinical Anatomy of the Cranial Nerves Clinical Anatomy of the Cranial Nerves
Clinical Anatomy of the Cranial Nerves Clinical Anatomy of the Cranial Nerves Paul Rea AMSTERDAM • BOSTON • HEIDELBERG • LONDON NEW YORK • OXFORD • PARIS • SAN DIEGO SAN FRANCISCO • SINGAPORE • SYDNEY • TOKYO Academic Press is an imprint of Elsevier Academic Press is an imprint of Elsevier 32 Jamestown Road, London NW1 7BY, UK The Boulevard, Langford Lane, Kidlington, Oxford OX5 1GB, UK Radarweg 29, PO Box 211, 1000 AE Amsterdam, The Netherlands 225 Wyman Street, Waltham, MA 02451, USA 525 B Street, Suite 1800, San Diego, CA 92101-4495, USA First published 2014 Copyright r 2014 Elsevier Inc. All rights reserved. No part of this publication may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, or any information storage and retrieval system, without permission in writing from the publisher. Details on how to seek permission, further information about the Publisher’s permissions policies and our arrangement with organizations such as the Copyright Clearance Center and the Copyright Licensing Agency, can be found at our website: www.elsevier.com/permissions. This book and the individual contributions contained in it are protected under copyright by the Publisher (other than as may be noted herein). Notices Knowledge and best practice in this field are constantly changing. As new research and experience broaden our understanding, changes in research methods, professional practices, or medical treatment may become necessary. Practitioners and researchers must always rely on their own experience and knowledge in evaluating and using any information, methods, compounds, or experiments described herein. In using such information or methods they should be mindful of their own safety and the safety of others, including parties for whom they have a professional responsibility. -

Quantifying Nerve Decussation Abnormalities in the Optic Chiasm T Robert J
NeuroImage: Clinical 24 (2019) 102055 Contents lists available at ScienceDirect NeuroImage: Clinical journal homepage: www.elsevier.com/locate/ynicl Quantifying nerve decussation abnormalities in the optic chiasm T Robert J. Puzniaka, Khazar Ahmadia, Jörn Kaufmannb, Andre Gouwsc, Antony B. Morlandc,d, ⁎ Franco Pestillie,1, Michael B. Hoffmanna,f,1, a Department of Ophthalmology, Otto-von-Guericke-University Magdeburg, Magdeburg, Germany b Department of Neurology, Otto-von-Guericke-University Magdeburg, Magdeburg, Germany c York Neuroimaging Centre, Department of Psychology, University of York, York, United Kingdom d York Biomedical Research Institute, University of York, York, United Kingdom e Department of Psychological and Brain Sciences, Program in Neuroscience and Program in Cognitive Science, Indiana University, Bloomington, USA f Center for Behavioral Brain Sciences, Magdeburg, Germany ARTICLE INFO ABSTRACT Keywords: Objective: The human optic chiasm comprises partially crossing optic nerve fibers. Here we used diffusion MRI Optic chiasm (dMRI) for the in-vivo identification of the abnormally high proportion of crossing fibers found intheoptic Albinism chiasm of people with albinism. Diffusion MRI Methods: In 9 individuals with albinism and 8 controls high-resolution 3T dMRI data was acquired and analyzed Functional MRI with a set of methods for signal modeling [Diffusion Tensor (DT) and Constrained Spherical Deconvolution Crossing nerves (CSD)], tractography, and streamline filtering (LiFE, COMMIT, and SIFT2). The number of crossing andnon- crossing streamlines and their weights after filtering entered ROC-analyses to compare the discriminative power of the methods based on the area under the curve (AUC). The dMRI results were cross-validated with fMRI estimates of misrouting in a subset of 6 albinotic individuals. -

The Irregular Chiasm C-Roughest Locus of Drosophila, Which Affects Axonal Projections and Programmed Cell Death: Encodes a Novel Immunoglobulin-Like Protein
Downloaded from genesdev.cshlp.org on October 9, 2021 - Published by Cold Spring Harbor Laboratory Press The irregular chiasm C-roughest locus of Drosophila, which affects axonal projections and programmed cell death: encodes a novel immunoglobulin-like protein Ricardo G.P. Ramos, 1 Gabor L. Igloi, Beate Lichte, Ute Baumann, 2 Dieter Maier, 3 Thilo Schneider, J. Helmut Brandst~itter, 4 Amalie Fr6hlich, s and Karl-Friedrich Fischbach 6 Institut f/Jr Biologie IU, Albert-Ludwigs Universit/it, D-79104 Freiburg im Breisgau, Germany; 3Biozentrum Basel, CH-4056 Basel, Switzerland The axonal projection mutations irregular chiasm C of Drosophila melanogaster comap and genetically interact with the roughest locus, which is required for programmed cell death in the developing retina. We cloned the genomic region in 3C5 by transposon tagging and identified a single transcription unit that produces a major, spatially and temporally regulated mRNA species of -5.0 kb. Postembryonic expression is strong in the developing optic lobe and in the eye imaginal disc. The gene encodes a transmembrane protein of 764 amino acids with five extracellular immunoglobulin-like domains and similarity to the chicken axonal surface glycoprotein DM-GRASP/SC1/BEN. Both known irreC alleles reduce the level of transcription, whereas the roughest cT mutation disrupts the intracellular domain of the protein. [Key Words: Cell adhesion; Immunoglobulin superfamily; DM-GRASP; optic chiasms; structural brain mutant; verticals] Received August 19, 1993; revised version accepted September 27, 1993. The assembly of a functional nervous system requires and extracellular matrix proteins, as well as the identi- the numerical matching and precise connection of neu- fication of factors, diffusible and localized, growth pro- ronal populations, which are often spatially distant.