Connective Tissue
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Basic Histology and Connective Tissue Chapter 5
Basic Histology and Connective Tissue Chapter 5 • Histology, the Study of Tissues • Tissue Types • Connective Tissues Histology is the Study of Tissues • 200 different types of cells in the human body. • A Tissue consist of two or more types of cells that function together. • Four basic types of tissues: – epithelial tissue – connective tissue – muscular tissue – nervous tissue • An Organ is a structure with discrete boundaries that is composed of 2 or more tissue types. • Example: skin is an organ composed of epidermal tissue and dermal tissue. Distinguishing Features of Tissue Types • Types of cells (shapes and functions) • Arrangement of cells • Characteristics of the Extracellular Matrix: – proportion of water – types of fibrous proteins – composition of the ground substance • ground substance is the gelatinous material between cells in addition to the water and fibrous proteins • ground substance consistency may be liquid (plasma), rubbery (cartilage), stony (bone), elastic (tendon) • Amount of space occupied by cells versus extracellular matrix distinguishes connective tissue from other tissues – cells of connective tissues are widely separated by a large amount of extracellular matrix – very little extracellular matrix between the cells of epithelia, nerve, and muscle tissue Embryonic Tissues • An embryo begins as a single cell that divides into many cells that eventually forms 3 Primary Layers: – ectoderm (outer layer) • forms epidermis and nervous system – endoderm (inner layer) • forms digestive glands and the mucous membrane lining digestive tract and respiratory system – mesoderm (middle layer) • Forms muscle, bone, blood and other organs. Histotechnology • Preparation of specimens for histology: – preserve tissue in a fixative to prevent decay (formalin) – dehydrate in solvents like alcohol and xylene – embed in wax or plastic – slice into very thin sections only 1 or 2 cells thick – float slices on water and mount on slides and then add color with stains • Sectioning an organ or tissue reduces a 3-dimensional structure to a 2- dimensional slice. -

Tendon Fibroblasts
Ann Rheum Dis: first published as 10.1136/ard.46.5.385 on 1 May 1987. Downloaded from Annals of the Rheumatic Diseases, 1987; 46, 385-390 Isolation and growth characteristics of adult human tendon fibroblasts M D CHARD, J K WRIGHT, AND B L HAZLEMAN From the Rheumatology Research Unit, Addenbrooke's Hospital, Hills Road, Cambridge SUMMARY An explant method for the isolation of fibroblasts from adult human tendon is described. Cells were successfully isolated from 22 out of 27 common biceps tendons obtained from cadaveric donors (age range 11-83 years). The fibroblasts could be maintained in culture using standard methods and morphologically resembled those of synovial rather than dermal origin. Growth characteristics of 12 cell lines were assessed by deoxyribose nucleic acid (DNA) synthesis using [3H]thymidine incorporation in response to stimulation by fetal calf serum. Cells obtained separately from superficial and deep parts of the tendons produced almost identical responses. No significant reduction in growth response with increasing age was found when related to the age of the donor. Therefore this study did not show any age related defect in the short term tendon fibroblast replicative responses to serum. Key words: in vitro, DNA synthesis. copyright. Tendinous lesions are common, but their nature and other tissues such as skin. It is desirable to investi- any possible predisposing factors are not well gate adult human tendon fibroblasts directly. It was understood. They occur in middle life frequently thought that the culture of adult human tendon after only minor, if any, injury. The extent to which fibroblasts would be very difficult to achieve be- this reflects age related changes in tendon tissue is cause of the differentiated appearance of cells on uncertain. -

Normal Gross and Histologic Features of the Gastrointestinal Tract
NORMAL GROSS AND HISTOLOGIC 1 FEATURES OF THE GASTROINTESTINAL TRACT THE NORMAL ESOPHAGUS left gastric, left phrenic, and left hepatic accessory arteries. Veins in the proximal and mid esopha- Anatomy gus drain into the systemic circulation, whereas Gross Anatomy. The adult esophagus is a the short gastric and left gastric veins of the muscular tube measuring approximately 25 cm portal system drain the distal esophagus. Linear and extending from the lower border of the cri- arrays of large caliber veins are unique to the distal coid cartilage to the gastroesophageal junction. esophagus and can be a helpful clue to the site of It lies posterior to the trachea and left atrium a biopsy when extensive cardiac-type mucosa is in the mediastinum but deviates slightly to the present near the gastroesophageal junction (4). left before descending to the diaphragm, where Lymphatic vessels are present in all layers of the it traverses the hiatus and enters the abdomen. esophagus. They drain to paratracheal and deep The subdiaphragmatic esophagus lies against cervical lymph nodes in the cervical esophagus, the posterior surface of the left hepatic lobe (1). bronchial and posterior mediastinal lymph nodes The International Classification of Diseases in the thoracic esophagus, and left gastric lymph and the American Joint Commission on Cancer nodes in the abdominal esophagus. divide the esophagus into upper, middle, and lower thirds, whereas endoscopists measure distance to points in the esophagus relative to the incisors (2). The esophagus begins 15 cm from the incisors and extends 40 cm from the incisors in the average adult (3). The upper and lower esophageal sphincters represent areas of increased resting tone but lack anatomic landmarks; they are located 15 to 18 cm from the incisors and slightly proximal to the gastroesophageal junction, respectively. -

The Plantar Aponeurosis in Fetuses and Adults: an Aponeurosis Or Fascia?
Int. J. Morphol., 35(2):684-690, 2017. The Plantar Aponeurosis in Fetuses and Adults: An Aponeurosis or Fascia? La Aponeurosis Plantar en Fetos y Adultos: ¿Aponeurosis o Fascia? A. Kalicharan; P. Pillay; C.O. Rennie; B.Z. De Gama & K.S. Satyapal KALICHARAN, A.; PILLAY, P.; RENNIE, C.O.; DE GAMA, B. Z. & SATYAPAL, K. S. The plantar aponeurosis in fetuses and adults: An aponeurosis or fascia? Int. J. Morphol., 35(2):684-690, 2017. SUMMARY: The plantar aponeurosis (PA), which is a thickened layer of deep fascia located on the plantar surface of the foot, is comprised of three parts. There are differing opinions on its nomenclature since various authors use the terms PA and plantar fascia (PF) interchangeably. In addition, the variable classifications of its parts has led to confusion. In order to assess the nature of the PA, this study documented its morphology. Furthermore, a pilot histological analysis was conducted to examine whether the structure is an aponeurosis or fascia. This study comprised of a morphological analysis of the three parts of the PA by micro- and macro-dissection of 50 fetal and 50 adult cadaveric feet, respectively (total n=100). Furthermore, a pilot histological analysis was conducted on five fetuses (n=10) and five adults (n=10) (total n=20). In each foot, the histological analysis was conducted on the three parts of the plantar aponeurosis, i.e. the central, lateral, and medial at their calcaneal origin (total n=60). Fetuses: i) Morphology: In 66 % (33/50) of the specimens, the standard anatomical pattern was observed, viz. -

The Mesocolon a Histological and Electron Microscopic Characterization of the Mesenteric Attachment of the Colon Prior to and After Surgical Mobilization
ORIGINAL ARTICLE The Mesocolon A Histological and Electron Microscopic Characterization of the Mesenteric Attachment of the Colon Prior to and After Surgical Mobilization Kevin Culligan, MRCS,∗ Stewart Walsh, FRCSEd,∗ Colum Dunne, PhD,∗ Michael Walsh, PhD,† Siobhan Ryan, MB,‡ Fabio Quondamatteo, MD,‡ Peter Dockery, PhD,§ and J. Calvin Coffey, FRCSI∗¶ uring fetal development, the dorsal mesentery suspends the en- Background: Colonic mobilization requires separation of mesocolon from tire gastrointestinal tract from the posterior abdominal wall. The underlying fascia. Despite the surgical importance of planes formed by these D mesocolon is the adult remnant of that part of the dorsal mesentery structures, no study has formally characterized their microscopic features. associated with the colon.1 In the adult human, the transverse and The aim of this study was to determine the histological and electron micro- lateral sigmoid portions of the mesocolon are mobile whereas the as- scopic appearance of mesocolon, fascia, and retroperitoneum, prior to and cending, descending, and medial sigmoid portions are nonmobile and after colonic mobilization. attached to underlying retroperitoneum.2–4 Classic anatomic teaching Methods: In 24 cadavers, samples were taken from right, transverse, de- maintains that the ascending and descending mesocolon “disappear” scending, and sigmoid mesocolon. In 12 cadavers, specimens were stained during embryogenesis.5,6 In keeping with this, the identification of a with hematoxylin and eosin (3 sections) or Masson trichrome (3 sections). In right or left mesocolon in the adult is frequently depicted as anoma- the second 12 cadavers, lymphatic channels were identified by staining im- lous rather than accepted as an anatomic norm.7 Accordingly, the munohistochemically for podoplanin. -

Characterization of Collagen Fibers (I, III, IV) and Elastin of Normal and Neoplastic Canine Prostatic Tissues
veterinary sciences Article Characterization of Collagen Fibers (I, III, IV) and Elastin of Normal and Neoplastic Canine Prostatic Tissues Luis Gabriel Rivera Calderón 1, Priscila Emiko Kobayashi 2, Rosemeri Oliveira Vasconcelos 1, Carlos Eduardo Fonseca-Alves 3 and Renée Laufer-Amorim 2,* 1 Department of Veterinary Pathology, School of Agricultural and Veterinarian Sciences, São Paulo State University (Unesp), Jaboticabal, São Paulo 14884-900, Brazil; [email protected] (L.G.R.C.); [email protected] (R.O.V.) 2 Department of Veterinary Clinic, School of Veterinary Medicine and Animal Science, São Paulo State University (Unesp), Botucatu, São Paulo 18618-681, Brazil; [email protected] 3 Department of Veterinary Surgery and Anesthesiology, School of Veterinary Medicine and Animal Science, São Paulo State University (Unesp), Botucatu, São Paulo 18618-681, Brazil; [email protected] * Correspondence: [email protected]; Tel.: +55-14-3880-2076 Received: 7 January 2019; Accepted: 25 February 2019; Published: 2 March 2019 Abstract: This study aimed to investigate collagen (Coll-I, III, IV) and elastin in canine normal prostate and prostate cancer (PC) using Picrosirius red (PSR) and Immunohistochemical (IHC) analysis. Eight normal prostates and 10 PC from formalin-fixed, paraffin-embedded samples were used. Collagen fibers area was analyzed with ImageJ software. The distribution of Coll-I and Coll-III was approximately 80% around prostatic ducts and acini, 15% among smooth muscle, and 5% surrounding blood vessels, in both normal prostate and PC. There was a higher median area of Coll-III in PC when compared to normal prostatic tissue (p = 0.001 for PSR and p = 0.05 for IHC). -

Nomina Histologica Veterinaria, First Edition
NOMINA HISTOLOGICA VETERINARIA Submitted by the International Committee on Veterinary Histological Nomenclature (ICVHN) to the World Association of Veterinary Anatomists Published on the website of the World Association of Veterinary Anatomists www.wava-amav.org 2017 CONTENTS Introduction i Principles of term construction in N.H.V. iii Cytologia – Cytology 1 Textus epithelialis – Epithelial tissue 10 Textus connectivus – Connective tissue 13 Sanguis et Lympha – Blood and Lymph 17 Textus muscularis – Muscle tissue 19 Textus nervosus – Nerve tissue 20 Splanchnologia – Viscera 23 Systema digestorium – Digestive system 24 Systema respiratorium – Respiratory system 32 Systema urinarium – Urinary system 35 Organa genitalia masculina – Male genital system 38 Organa genitalia feminina – Female genital system 42 Systema endocrinum – Endocrine system 45 Systema cardiovasculare et lymphaticum [Angiologia] – Cardiovascular and lymphatic system 47 Systema nervosum – Nervous system 52 Receptores sensorii et Organa sensuum – Sensory receptors and Sense organs 58 Integumentum – Integument 64 INTRODUCTION The preparations leading to the publication of the present first edition of the Nomina Histologica Veterinaria has a long history spanning more than 50 years. Under the auspices of the World Association of Veterinary Anatomists (W.A.V.A.), the International Committee on Veterinary Anatomical Nomenclature (I.C.V.A.N.) appointed in Giessen, 1965, a Subcommittee on Histology and Embryology which started a working relation with the Subcommittee on Histology of the former International Anatomical Nomenclature Committee. In Mexico City, 1971, this Subcommittee presented a document entitled Nomina Histologica Veterinaria: A Working Draft as a basis for the continued work of the newly-appointed Subcommittee on Histological Nomenclature. This resulted in the editing of the Nomina Histologica Veterinaria: A Working Draft II (Toulouse, 1974), followed by preparations for publication of a Nomina Histologica Veterinaria. -
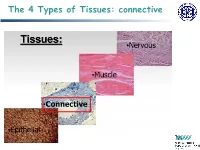
The 4 Types of Tissues: Connective
The 4 Types of Tissues: connective Connective Tissue General structure of CT cells are dispersed in a matrix matrix = a large amount of extracellular material produced by the CT cells and plays a major role in the functioning matrix component = ground substance often crisscrossed by protein fibers ground substance usually fluid, but it can also be mineralized and solid (bones) CTs = vast variety of forms, but typically 3 characteristic components: cells, large amounts of amorphous ground substance, and protein fibers. Connective Tissue GROUND SUBSTANCE In connective tissue, the ground substance is an amorphous gel-like substance surrounding the cells. In a tissue, cells are surrounded and supported by an extracellular matrix. Ground substance traditionally does not include fibers (collagen and elastic fibers), but does include all the other components of the extracellular matrix . The components of the ground substance vary depending on the tissue. Ground substance is primarily composed of water, glycosaminoglycans (most notably hyaluronan ), proteoglycans, and glycoproteins. Usually it is not visible on slides, because it is lost during the preparation process. Connective Tissue Functions of Connective Tissues Support and connect other tissues Protection (fibrous capsules and bones that protect delicate organs and, of course, the skeletal system). Transport of fluid, nutrients, waste, and chemical messengers is ensured by specialized fluid connective tissues, such as blood and lymph. Adipose cells store surplus energy in the form of fat and contribute to the thermal insulation of the body. Embryonic Connective Tissue All connective tissues derive from the mesodermal layer of the embryo . The first connective tissue to develop in the embryo is mesenchyme , the stem cell line from which all connective tissues are later derived. -

Brown Adipose Tissue: New Challenges for Prevention of Childhood Obesity
nutrients Review Brown Adipose Tissue: New Challenges for Prevention of Childhood Obesity. A Narrative Review Elvira Verduci 1,2,*,† , Valeria Calcaterra 2,3,† , Elisabetta Di Profio 2,4, Giulia Fiore 2, Federica Rey 5,6 , Vittoria Carlotta Magenes 2, Carolina Federica Todisco 2, Stephana Carelli 5,6,* and Gian Vincenzo Zuccotti 2,5,6 1 Department of Health Sciences, University of Milan, 20146 Milan, Italy 2 Department of Pediatrics, Vittore Buzzi Children’s Hospital, University of Milan, 20154 Milan, Italy; [email protected] (V.C.); elisabetta.diprofi[email protected] (E.D.P.); giulia.fi[email protected] (G.F.); [email protected] (V.C.M.); [email protected] (C.F.T.); [email protected] (G.V.Z.) 3 Pediatric and Adolescent Unit, Department of Internal Medicine, University of Pavia, 27100 Pavia, Italy 4 Department of Animal Sciences for Health, Animal Production and Food Safety, University of Milan, 20133 Milan, Italy 5 Department of Biomedical and Clinical Sciences “L. Sacco”, University of Milan, 20157 Milan, Italy; [email protected] 6 Pediatric Clinical Research Center Fondazione Romeo ed Enrica Invernizzi, University of Milan, 20157 Milan, Italy * Correspondence: [email protected] (E.V.); [email protected] (S.C.) † These authors contributed equally to this work. Abstract: Pediatric obesity remains a challenge in modern society. Recently, research has focused on the role of the brown adipose tissue (BAT) as a potential target of intervention. In this review, we Citation: Verduci, E.; Calcaterra, V.; revised preclinical and clinical works on factors that may promote BAT or browning of white adipose Di Profio, E.; Fiore, G.; Rey, F.; tissue (WAT) from fetal age to adolescence. -

Histology of Watersnake (Enhydris Enhydris) Digestive System
E3S Web of Conferences 151, 01052 (2020) https://doi.org/10.1051/e3sconf/202015101052 st 1 ICVAES 2019 Histology of Watersnake (Enhydris Enhydris) Digestive System Dian Masyitha1,*, Lena Maulidar 2 , Zainuddin Zainuddin1, Muhammad N. Salim3, Dwinna Aliza3, Fadli A. Gani 4 , Rusli Rusli5 1 Histology Laboratory of the Faculty of Veterinary Medicine, Syiah Kuala University, Banda Aceh 23111, Indonesia 2 Veterinary Education Study Program Faculty of Veterinary Medicine, Syiah Kuala University, Banda Aceh 23111, Indonesia 3 Pathology Laboratory, Faculty of Veterinary Medicine, Universitas Syiah Kuala , Banda Aceh, Indonesia 4 Anatomy Laboratory, Faculty of Veterinary Medicine, Universitas Syiah Kuala , Banda Aceh, Indonesia 5 Clinical Laboratory of the Faculty of Veterinary Medicine, Universitas Syiah Kuala , Banda Aceh, Indonesia Abstract. This research aimed to study the histology of the digestive system of the watersnake (Enhydris enhydris). The digestive system taken was the esophagus, stomach, frontal small intestine and the back of the large intestine from three watersnakes. The samples were then made into histological preparations with hematoxylin-eosin (HE) staining and observed exploratively. The results showed that the digestive system of the watersnake was composed of layers of tissue, namely the mucosa, tunica submucosa, tunica muscularis, and serous tunica. Mucosal mucosa consisted of the lamina epithelium, lamina propria, and mucous lamina muscularis. The submucosal tunica consisted of connective tissue with blood vessels, lymph, and nerves. The muscular tunica was composed of circular muscles and elongated muscles. The serous tunica consisted of a thin layer of connective tissue that was covered by a thin layer of the mesothelium (mesothelium). The histological structure of the snake digestive system is not much different from the reptile digestive system. -

Collagen Type I and Decorin Expression in Tenocytes Depend On
Wagenhäuser et al. BMC Musculoskeletal Disorders 2012, 13:140 http://www.biomedcentral.com/1471-2474/13/140 RESEARCH ARTICLE Open Access Collagen type I and decorin expression in tenocytes depend on the cell isolation method Markus U Wagenhäuser1†, Matthias F Pietschmann1*†, Birte Sievers1, Denitsa Docheva2, Matthias Schieker2, Volkmar Jansson1 and Peter E Müller1 Abstract Backround: The treatment of rotator cuff tears is still challenging. Tendon tissue engineering (TTE) might be an alternative in future. Tenocytes seem to be the most suitable cell type as they are easy to obtain and no differentiation in vitro is necessary. The aim of this study was to examine, if the long head of the biceps tendon (LHB) can deliver viable tenocytes for TTE. In this context, different isolation methods, such as enzymatic digestion (ED) and cell migration (CM), are investigated on differences in gene expression and cell morphology. Methods: Samples of the LHB were obtained from patients, who underwent surgery for primary shoulder arthroplasty. Using ED as isolation method, 0.2% collagenase I solution was used. Using CM as isolation method, small pieces of minced tendon were put into petri-dishes. After cell cultivation, RT-PCR was performed for collagen type I, collagen type III, decorin, tenascin-C, fibronectin, Scleraxis, tenomodulin, osteopontin and agreccan. Results: The total number of isolated cells, in relation to 1 g of native tissue, was 14 times higher using ED. The time interval for cell isolation was about 17 hours using ED and approximately 50 days using CM. Cell morphology in vitro was similar for both isolation techniques. Higher expression of collagen type I could be observed in tenocyte-like cell cultures (TLCC) using ED as isolation method (p < 0.05), however decorin expression was higher in TLCC using CM as isolation method (p < 0.05). -

Autoimmunity Mixed Connective Tissue Disease (CTD)
Autoimmunity Mixed Connective Tissue Disease (mixed CTD) and Undifferentiated Connective Tissue Disease (UCTD) Autoimmunity and Connective Tissue Disease (CTD) The immune system normally produces antibodies which attack bugs (viruses, bacteria and fungi). Sometimes, for reasons we don’t fully understand, the immune system goes into ‘overdrive’ and produces antibodies which attack the body’s own tissues, causing inflammation. This is called autoimmunity and may cause an autoimmune disease. A common example of this is underactive thyroid where antibodies are produced which attack the thyroid gland. The connective tissues are the structural portions of our body that essentially hold the cells of the body together. These tissues form a framework or matrix for the body. Connective Tissue Disease (CTD) Connective tissue disease is an autoimmune disease where the body produces antibodies against its own connective tissue, causing inflammation. The ‘classic’ connective tissue diseases include: Lupus Rheumatoid arthritis Scleroderma (or systemic sclerosis) Polymyositis and Source: Rheumatology Reference No: 6252-1 Issue date: 26/9/19 Review date: 26/9/22 Page 1 of 4 Dermatomyositis Each of these diseases has a typical presentation with clinical findings that doctors can recognise during an examination. Each also has certain blood test abnormalities and abnormal antibody patterns. However, each of these diseases can start with very mild symptoms before developing the classic features that help in the diagnosis. Undifferentiated Connective Tissue Disease (UCTD) Almost one in four people seen in rheumatology clinics develop an autoimmune disease which doesn't fit neatly into a category, so they are not given a definite disease label. When these conditions have not developed the classic features of a particular disease, doctors will often refer to the condition as "undifferentiated connective tissue disease" or UCTD for short.