Passive Electrical Properties of the Neuron
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Servo Motor EMMT-AS-60-L-LS-RM Part Number: 5242213
Servo motor EMMT-AS-60-L-LS-RM Part number: 5242213 Data sheet Feature Value Short type code EMMT-AS Ambient temperature -15 °C ... 40 °C Note on ambient temperature Up to 80°C with derating of -1.5% per degree Celsius Max. installation height 4000 m Note on max. installation height As of 1,000 m: only with derating of -1.0% per 100 m Storage temperature -20 °C ... 70 °C Relative air humidity 0 - 90% Conforms to standard IEC 60034 Temperature class as per EN 60034-1 F Max. winding temperature 155 °C Rating class as per EN 60034-1 S1 Temperature monitoring Digital motor temperature transmission via EnDat® 2.2 Motor type to EN 60034-7 IM B5 IM V1 IM V3 Mounting position optional Degree of protection IP40 Note on degree of protection IP40 for motor shaft without rotary shaft seal IP65 for motor shaft with rotary shaft seal IP67 for motor housing including connection components Concentricity, coaxiality, axial runout to DIN SPEC 42955 N Balance quality G 2.5 Detent torque <1.0% of the peak torque Storage lifetime under nominal conditions 20000 h Interface code, motor out 60P Electrical connection 1, connection type Hybrid plug Electrical connection 1, connector system M23x1 Electrical connection 1, number of connections/cores 15 Electrical connection 1, connection pattern 00995913 Pollution degree 2 Note on materials RoHS-compliant Corrosion resistance class CRC 0 - No corrosion stress LABS-Conformity VDMA24364 zone III Vibration resistance Transport application test with severity level 2 to FN 942017-4 and EN 60068-2-6 10/2/21 - Subject to change - Festo AG & Co. -
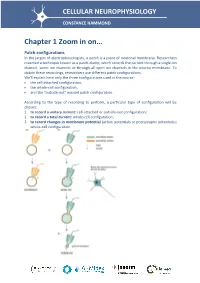
Chapter 1 Zoom in On… Patch Configurations in the Jargon of Electrophysiologists, a Patch Is a Piece of Neuronal Membrane
CELLULAR NEUROPHYSIOLOGY CONSTANCE HAMMOND Chapter 1 Zoom in on… Patch configurations In the jargon of electrophysiologists, a patch is a piece of neuronal membrane. Researchers invented a technique known as a patch-clamp, which records the current through a single ion channel, some ion channels or through all open ion channels in the neuron membrane. To obtain these recordings, researchers use different patch configurations. We'll explain here only the three configurations used in the course: the cell-attached configuration, the whole-cell configuration, and the "outside-out" excised patch configuration. According to the type of recording to perform, a particular type of configuration will be chosen: 1. to record a unitary current: cell-attached or outside-out configuration; 2. to record a total current: whole-cell configuration; 3. to record changes in membrane potential (action potentials or postsynaptic potentials): whole-cell configuration. The cell-attached configuration (attached to the pipette - Figure a) First, the pipette is filled with an extracellular fluid. Positive pressure is applied in the electrode by means of a syringe connected to the electrode, so that the intrapipette fluid tends to leave the pipette. The electrode is then brought near to the soma of a neuron. When the electrode touches the membrane, the positive pressure is withdrawn to draw the membrane toward the mouth of the pipette. We wait without moving the pipette; a seal is made between the walls of the pipette and the soma membrane. This seal strength can be measured by applying a low level of amplitude current in the pipette. Since V = RI, one can measure the resistance of the seal. -

Crayfish Escape Behavior and Central Synapses
Crayfish Escape Behavior and Central Synapses. III. Electrical Junctions and Dendrite Spikes in Fast Flexor Motoneurons ROBERT S. ZUCKER Department of Biological Sciences and Program in the Neurological Sciences, Stanford University, Stanford, California 94305 THE LATERAL giant fiber is the decision fiber and fast flexor motoneurons are located in for a behavioral response in crayfish in- the dorsal neuropil of each abdominal gan- volving rapid tail flexion in response to glion. Dye injections and anatomical recon- abdominal tactile stimulation (27). Each structions of ‘identified motoneurons reveal lateral giant impulse is followed by a rapid that only those motoneurons which have tail flexion or tail flip, and when the giant dentritic branches in contact with a giant fiber does not fire in response to such stim- fiber are activated by that giant fiber (12, uli, the movement does not occur. 21). All of tl ie fast flexor motoneurons are The afferent limb of the reflex exciting excited by the ipsilateral giant; all but F4, the lateral giant has been described (28), F5 and F7 are also excited by the contra- and the habituation of the behavior has latkral giant (21 a). been explained in terms of the properties The excitation of these motoneurons, of this part of the neural circuit (29). seen as depolarizing potentials in their The properties of the neuromuscular somata, does not always reach threshold for junctions between the fast flexor motoneu- generating a spike which propagates out the rons and the phasic flexor muscles have also axon to the periphery (14). Indeed, only t,he been described (13). -

Long-Term Adult Human Brain Slice Cultures As a Model System to Study
TOOLS AND RESOURCES Long-term adult human brain slice cultures as a model system to study human CNS circuitry and disease Niklas Schwarz1, Betu¨ l Uysal1, Marc Welzer2,3, Jacqueline C Bahr1, Nikolas Layer1, Heidi Lo¨ ffler1, Kornelijus Stanaitis1, Harshad PA1, Yvonne G Weber1,4, Ulrike BS Hedrich1, Ju¨ rgen B Honegger4, Angelos Skodras2,3, Albert J Becker5, Thomas V Wuttke1,4†*, Henner Koch1†* 1Department of Neurology and Epileptology, Hertie-Institute for Clinical Brain Research, University of Tu¨ bingen, Tu¨ bingen, Germany; 2Department of Cellular Neurology, Hertie-Institute for Clinical Brain Research, University of Tu¨ bingen, Tu¨ bingen, Germany; 3German Center for Neurodegenerative Diseases (DZNE), Tu¨ bingen, Germany; 4Department of Neurosurgery, University of Tu¨ bingen, Tu¨ bingen, Germany; 5Department of Neuropathology, Section for Translational Epilepsy Research, University Bonn Medical Center, Bonn, Germany Abstract Most of our knowledge on human CNS circuitry and related disorders originates from model organisms. How well such data translate to the human CNS remains largely to be determined. Human brain slice cultures derived from neurosurgical resections may offer novel avenues to approach this translational gap. We now demonstrate robust preservation of the complex neuronal cytoarchitecture and electrophysiological properties of human pyramidal neurons *For correspondence: in long-term brain slice cultures. Further experiments delineate the optimal conditions for efficient [email protected] viral transduction of cultures, enabling ‘high throughput’ fluorescence-mediated 3D reconstruction tuebingen.de (TVW); of genetically targeted neurons at comparable quality to state-of-the-art biocytin fillings, and [email protected] demonstrate feasibility of long term live cell imaging of human cells in vitro. -

Nernst Potentials and Membrane Potential Changes
UNDERSTANDING MEMBRANE POTENTIAL CHANGES IN TERMS OF NERNST POTENTIALS: For seeing how a change in conductance to ions affects the membrane potential, follow these steps: 1. Make a graph with membrane potential on the vertical axis (-100 to +55) and time on the horizontal axis. 2. Draw dashed lines indicating the standard Nernst potential (equilibrium potential) for each ion: Na+ = +55 mV, K+ = -90mV, Cl- = -65 mV. 3. Draw lines below the horizontal axis showing the increased conductance to individual ions. 4. Start plotting the membrane potential on the left. Most graphs will start at resting potential (-70 mV) 5. When current injection (Stim) is present, move the membrane potential upward to Firing threshold. 6. For the time during which membrane conductance to a particular ion increases, move the membrane potential toward the Nernst potential for that ion. 7. During the time when conductance to a particular ion decreases, move the membrane potential away from the Nernst potential of that ion, toward a position which averages the conductances of the other ions. 8. When conductances return to their original value, membrane potential will go to its starting value. +55mV ACTION POTENTIAL SYNAPTIC POTENTIALS 0mV Membrane potential Firing threshold Firing threshold -65mV -90mV Time Time Na+ K+ Conductances Stim CHANGES IN MEMBRANE POTENTIAL ALLOW NEURONS TO COMMUNICATE The membrane potential of a neuron can be measured with an intracellular electrode. This 1 provides a measurement of the voltage difference between the inside of the cell and the outside. When there is no external input, the membrane potential will usually remain at a value called the resting potential. -

Unix Programming
P.G DEPARTMENT OF COMPUTER APPLICATIONS 18PMC532 UNIX PROGRAMMING K1 QUESTIONS WITH ANSWERS UNIT- 1 1) Define unix. Unix was originally written in assembler, but it was rewritten in 1973 in c, which was principally authored by Dennis Ritchie ( c is based on the b language developed by kenThompson. 2) Discuss the Communication. Excellent communication with users, network User can easily exchange mail,dta,pgms in the network 3) Discuss Security Login Names Passwords Access Rights File Level (R W X) File Encryption 4) Define PORTABILITY UNIX run on any type of Hardware and configuration Flexibility credits goes to Dennis Ritchie( c pgms) Ported with IBM PC to GRAY 2 5) Define OPEN SYSTEM Everything in unix is treated as file(source pgm, Floppy disk,printer, terminal etc., Modification of the system is easy because the Source code is always available 6) The file system breaks the disk in to four segements The boot block The super block The Inode table Data block 7) Command used to find out the block size on your file $cmchk BSIZE=1024 8) Define Boot Block Generally the first block number 0 is called the BOOT BLOCK. It consists of Hardware specific boot program that loads the file known as kernal of the system. 9) Define super block It describes the state of the file system ie how large it is and how many maximum Files can it accommodate This is the 2nd block and is number 1 used to control the allocation of disk blocks 10) Define inode table The third segment includes block number 2 to n of the file system is called Inode Table. -

Useful Commands in Linux and Other Tools for Quality Control
Useful commands in Linux and other tools for quality control Ignacio Aguilar INIA Uruguay 05-2018 Unix Basic Commands pwd show working directory ls list files in working directory ll as before but with more information mkdir d make a directory d cd d change to directory d Copy and moving commands To copy file cp /home/user/is . To copy file directory cp –r /home/folder . to move file aa into bb in folder test mv aa ./test/bb To delete rm yy delete the file yy rm –r xx delete the folder xx Redirections & pipe Redirection useful to read/write from file !! aa < bb program aa reads from file bb blupf90 < in aa > bb program aa write in file bb blupf90 < in > log Redirections & pipe “|” similar to redirection but instead to write to a file, passes content as input to other command tee copy standard input to standard output and save in a file echo copy stream to standard output Example: program blupf90 reads name of parameter file and writes output in terminal and in file log echo par.b90 | blupf90 | tee blup.log Other popular commands head file print first 10 lines list file page-by-page tail file print last 10 lines less file list file line-by-line or page-by-page wc –l file count lines grep text file find lines that contains text cat file1 fiel2 concatenate files sort sort file cut cuts specific columns join join lines of two files on specific columns paste paste lines of two file expand replace TAB with spaces uniq retain unique lines on a sorted file head / tail $ head pedigree.txt 1 0 0 2 0 0 3 0 0 4 0 0 5 0 0 6 0 0 7 0 0 8 0 0 9 0 0 10 -

The Narrowing of Dendrite Branches Across Nodes Follows a Well-Defined Scaling Law
The narrowing of dendrite branches across nodes follows a well-defined scaling law Maijia Liaoa, Xin Liangb, and Jonathon Howarda,1 aDepartment of Molecular Biophysics and Biochemistry, Yale University, New Haven, CT 06520; and bTsinghua-Peking Joint Center for Life Sciences, School of Life Sciences, Tsinghua University, 100084 Beijing, China Edited by Yuh Nung Jan, Howard Hughes Medical Institute, University of California San Francisco, San Francisco, CA, and approved May 17, 2021 (received for review October 28, 2020) The systematic variation of diameters in branched networks has minimized. Da Vinci’s law has been reformulated as the “pipe tantalized biologists since the discovery of da Vinci’s rule for trees. model” for plants, in which a fixed cross-section of stems and Da Vinci’s rule can be formulated as a power law with exponent branches is required to support each unit amount of leaves (13). two: The square of the mother branch’s diameter is equal to the Experimental support for scaling laws, however, is scarce because sum of the squares of those of the daughters. Power laws, with of the difficulties of imaging entire branched networks and because different exponents, have been proposed for branching in circula- intrinsic anatomical variability may obscure precise laws (14). Thus, tory systems (Murray’s law with exponent 3) and in neurons (Rall’s it is an open question whether scaling laws such as Eq. 1 apply in law with exponent 3/2). The laws have been derived theoretically, biological systems. based on optimality arguments, but, for the most part, have not To quantitatively test diameter-scaling laws in neurons, it is been tested rigorously. -

Effect of Hyperkalemia on Membrane Potential: Depolarization
❖ CASE 3 A 6-year-old boy is brought to the family physician after his parents noticed that he had difficulty moving his arms and legs after a soccer game. About 10 minutes after leaving the field, the boy became so weak that he could not stand for about 30 minutes. Questioning revealed that he had complained of weakness after eating bananas, had frequent muscle spasms, and occasionally had myotonia, which was expressed as difficulty in releasing his grip or diffi- culty opening his eyes after squinting into the sun. After a thorough physical examination, the boy was diagnosed with hyperkalemic periodic paralysis. The family was advised to feed the boy carbohydrate-rich, low-potassium foods, give him glucose-containing drinks during attacks, and have him avoid strenuous exercise and fasting. ◆ What is the effect of hyperkalemia on cell membrane potential? ◆ What is responsible for the repolarizing phase of an action potential? ◆ What is the effect of prolonged depolarization on the skeletal muscle Na+ channel? 32 CASE FILES: PHYSIOLOGY ANSWERS TO CASE 3: ACTION POTENTIAL Summary: A 6-year-old boy who experiences profound weakness after exer- cise is diagnosed with hyperkalemic periodic paralysis. ◆ Effect of hyperkalemia on membrane potential: Depolarization. ◆ Repolarization mechanisms: Activation of voltage-gated K+ conductance and inactivation of Na+ conductance. ◆ Effect of prolonged depolarization: Inactivation of Na+ channels. CLINICAL CORRELATION Hyperkalemic periodic paralysis (HyperPP) is a dominant inherited trait caused by a mutation in the α subunit of the skeletal muscle Na+ channel. It occurs in approximately 1 in 100,000 people and is more common and more severe in males. -

Configuring Your Login Session
SSCC Pub.# 7-9 Last revised: 5/18/99 Configuring Your Login Session When you log into UNIX, you are running a program called a shell. The shell is the program that provides you with the prompt and that submits to the computer commands that you type on the command line. This shell is highly configurable. It has already been partially configured for you, but it is possible to change the way that the shell runs. Many shells run under UNIX. The shell that SSCC users use by default is called the tcsh, pronounced "Tee-Cee-shell", or more simply, the C shell. The C shell can be configured using three files called .login, .cshrc, and .logout, which reside in your home directory. Also, many other programs can be configured using the C shell's configuration files. Below are sample configuration files for the C shell and explanations of the commands contained within these files. As you find commands that you would like to include in your configuration files, use an editor (such as EMACS or nuTPU) to add the lines to your own configuration files. Since the first character of configuration files is a dot ("."), the files are called "dot files". They are also called "hidden files" because you cannot see them when you type the ls command. They can only be listed when using the -a option with the ls command. Other commands may have their own setup files. These files almost always begin with a dot and often end with the letters "rc", which stands for "run commands". -

Electrical Activity of the Heart: Action Potential, Automaticity, and Conduction 1 & 2 Clive M
Electrical Activity of the Heart: Action Potential, Automaticity, and Conduction 1 & 2 Clive M. Baumgarten, Ph.D. OBJECTIVES: 1. Describe the basic characteristics of cardiac electrical activity and the spread of the action potential through the heart 2. Compare the characteristics of action potentials in different parts of the heart 3. Describe how serum K modulates resting potential 4. Describe the ionic basis for the cardiac action potential and changes in ion currents during each phase of the action potential 5. Identify differences in electrical activity across the tissues of the heart 6. Describe the basis for normal automaticity 7. Describe the basis for excitability 8. Describe the basis for conduction of the cardiac action potential 9. Describe how the responsiveness relationship and the Na+ channel cycle modulate cardiac electrical activity I. BASIC ELECTROPHYSIOLOGIC CHARACTERISTICS OF CARDIAC MUSCLE A. Electrical activity is myogenic, i.e., it originates in the heart. The heart is an electrical syncitium (i.e., behaves as if one cell). The action potential spreads from cell-to-cell initiating contraction. Cardiac electrical activity is modulated by the autonomic nervous system. B. Cardiac cells are electrically coupled by low resistance conducting pathways gap junctions located at the intercalated disc, at the ends of cells, and at nexus, points of side-to-side contact. The low resistance pathways (wide channels) are formed by connexins. Connexins permit the flow of current and the spread of the action potential from cell-to-cell. C. Action potentials are much longer in duration in cardiac muscle (up to 400 msec) than in nerve or skeletal muscle (~5 msec). -

NEURAL CONNECTIONS: Some You Use, Some You Lose
NEURAL CONNECTIONS: Some You Use, Some You Lose by JOHN T. BRUER SOURCE: Phi Delta Kappan 81 no4 264-77 D 1999 . The magazine publisher is the copyright holder of this article and it is reproduced with permission. Further reproduction of this article in violation of the copyright is prohibited JOHN T. BRUER is president of the James S. McDonnell Foundation, St. Louis. This article is adapted from his new book, The Myth of the First Three Years (Free Press, 1999), and is reprinted by arrangement with The Free Press, a division of Simon Schuster Inc. ©1999, John T. Bruer . OVER 20 YEARS AGO, neuroscientists discovered that humans and other animals experience a rapid increase in brain connectivity -- an exuberant burst of synapse formation -- early in development. They have studied this process most carefully in the brain's outer layer, or cortex, which is essentially our gray matter. In these studies, neuroscientists have documented that over our life spans the number of synapses per unit area or unit volume of cortical tissue changes, as does the number of synapses per neuron. Neuroscientists refer to the number of synapses per unit of cortical tissue as the brain's synaptic density. Over our lifetimes, our brain's synaptic density changes in an interesting, patterned way. This pattern of synaptic change and what it might mean is the first neurobiological strand of the Myth of the First Three Years. (The second strand of the Myth deals with the notion of critical periods, and the third takes up the matter of enriched, or complex, environments.) Popular discussions of the new brain science trade heavily on what happens to synapses during infancy and childhood.