The Influence of Longitudinal Hydrological Connectivity on Resource Availability and Lower Order Food Web Structure in the Murray River
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

NORTH WEST Freight Transport Strategy
NORTH WEST Freight Transport Strategy Department of Infrastructure NORTH WEST FREIGHT TRANSPORT STRATEGY Final Report May 2002 This report has been prepared by the Department of Infrastructure, VicRoads, Mildura Rural City Council, Swan Hill Rural City Council and the North West Municipalities Association to guide planning and development of the freight transport network in the north-west of Victoria. The State Government acknowledges the participation and support of the Councils of the north-west in preparing the strategy and the many stakeholders and individuals who contributed comments and ideas. Department of Infrastructure Strategic Planning Division Level 23, 80 Collins St Melbourne VIC 3000 www.doi.vic.gov.au Final Report North West Freight Transport Strategy Table of Contents Executive Summary ......................................................................................................................... i 1. Strategy Outline. ...........................................................................................................................1 1.1 Background .............................................................................................................................1 1.2 Strategy Outcomes.................................................................................................................1 1.3 Planning Horizon.....................................................................................................................1 1.4 Other Investigations ................................................................................................................1 -

Bridge Types in NSW Historical Overviews 2006
Bridge Types in NSW Historical overviews 2006 These historical overviews of bridge types in NSW are extracts compiled from bridge population studies commissioned by RTA Environment Branch. CONTENTS Section Page 1. Masonry Bridges 1 2. Timber Beam Bridges 12 3. Timber Truss Bridges 25 4. Pre-1930 Metal Bridges 57 5. Concrete Beam Bridges 75 6. Concrete Slab and Arch Bridges 101 Masonry Bridges Heritage Study of Masonry Bridges in NSW 2005 1 Historical Overview of Bridge Types in NSW: Extract from the Study of Masonry Bridges in NSW HISTORICAL BACKGROUND TO MASONRY BRIDGES IN NSW 1.1 History of early bridges constructed in NSW Bridges constructed prior to the 1830s were relatively simple forms. The majority of these were timber structures, with the occasional use of stone piers. The first bridge constructed in NSW was built in 1788. The bridge was a simple timber bridge constructed over the Tank Stream, near what is today the intersection of George and Bridge Streets in the Central Business District of Sydney. Soon after it was washed away and needed to be replaced. The first "permanent" bridge in NSW was this bridge's successor. This was a masonry and timber arch bridge with a span of 24 feet erected in 1803 (Figure 1.1). However this was not a triumph of colonial bridge engineering, as it collapsed after only three years' service. It took a further five years for the bridge to be rebuilt in an improved form. The contractor who undertook this work received payment of 660 gallons of spirits, this being an alternative currency in the Colony at the time (Main Roads, 1950: 37) Figure 1.1 “View of Sydney from The Rocks, 1803”, by John Lancashire (Dixson Galleries, SLNSW). -

SPECIAL Victoria Government Gazette
Victoria Government Gazette No. S 6 Wednesday 6 January 2010 By Authority of Victorian Government Printer Road Safety Act 1986 ROAD SAFETY (VEHICLES) REGULATIONS 2009 Specifi cation of Road Train Routes 1. Purpose The purpose of this notice is to specify road train routes for the purposes of the Road Safety (Vehicles) Regulations 2009 (in this notice referred to as the Vehicles Regulations). 2. Authorising provisions This notice is made under clauses 21 and 31 of Schedule 8 to the Vehicles Regulations. Clause 21 of Schedule 8 to the Vehicles Regulations provides that a road train may only be used on a highway in an area, or on a route, specifi ed by the Roads Corporation by notice published in the Government Gazette. Clause 31 of Schedule 8 to the Vehicles Regulations provides that a prime mover, semi-trailer and converter dolly combination may only be used on a highway in an area, or on a route, specifi ed by the Roads Corporation by notice published in the Government Gazette. 3. Commencement This notice comes into operation on the date it is published in the Government Gazette. 4. Revocation The notice published in Government Gazette No. S 200 on Tuesday 25 October 2005 entitled Notice Concerning Road Train Routes is revoked. 5. Expiry This notice expires on 9 November 2014. 6. Specifi cation For the purposes of clauses 21 and 31 of Schedule 8 to the Vehicles Regulations, I, George Mavroyeni, delegate of the Roads Corporation, specify the following Victorian routes as ones on which road trains may be used, subject to satisfying the requirements -

Patricia Palmer Lee PRG 1722 Special List POSTCARDS INDEX
___________________________________________________________ ______________________ Patricia Palmer Lee PRG 1722 Special List POSTCARDS INDEX 1993 to 2014 NO. DATE SUBJECT POSTMARK STAMPS A1 05.07.1993 Ramsgate Beach, Botany Bay Sydney Parma Wallaby A2 09.07.1993 Bondi Beach Surf Eastern Suburbs Ghost Bat A3 13.07.1993 Autumn Foliage, Blue Mountains Eastern Suburbs Tasmanian Herit Train A4 20.07.1993 Baha'i Temple, Ingleside Eastern Suburbs Silver City Comet A5 27.07.1993 Harbour Bridge from McMahon's Point Eastern Suburbs Kuranda Tourist Train A6 04.08.1993 Winter Sunset, Cooks River, Tempe Eastern Suburbs Long-tailed Dunnart A7 10.08.1993 Henry Lawson Memorial, Domain Eastern Suburbs Little Pygmy-Possum A8 17.08.1993 Berry Island, Parramatta River Rushcutters Bay Ghost Bat A9 24.08.1993 Story Bridge, Brisbane River Eastern Suburbs Parma Wallaby A10 28.08.1993 Stradbroke Island, Moreton Bay Qld Cootamundra Long-tailed Dunnart A11 31.08.1993 Rainforest, Brisbane Botanical Gardens Yass Little Pygmy-Possum A12 05.09.1993 Dinosaur Exhibit, Brisbane Museum Eastern Suburbs Ghost Bat A13 10.09.1993 Wattle Festival Time, Cootamundra Eastern Suburbs Squirrel Glider A14 14.09.1993 Davidson Nat Park, Middle Harbour Eastern Suburbs Dusky Hopping-Mouse A15 17.09.1993 Cooma Cottage, Yass Eastern Suburbs Parma Wallaby A16 21.09.1993 Bicentennial Park, Homebush Bay Eastern Suburbs The Ghan A17 24.09.1993 Rainbow, North Coast NSW Eastern Suburbs Long-Tailed Dunnart A18 28.09.1993 Sphinx Monument, Kuring-gai Chase NP Canberra Little Pygmy-Possum A19 01.10.1993 -

Transport for NSW Faqs 25 OCTOBER 2019
Transport for NSW FAQs 25 OCTOBER 2019 Emergency closure of Bunnerungee Bridge Why is the highway closed at Bunnerungee Bridge? A structural problem has been identified following a routine bridge inspection at Bunnerungee Bridge, 65 kilometres north of Wentworth on the Silver City Highway. How long is the bridge expected to be closed? A specialist Road and Maritime Services/ Transport for NSW bridge team is investigating and assessing the bridge while working on a solution. It is not yet known how long the repairs will take. Roads and Maritime Services will open the bridge as soon as it is safe to do so. The community will be kept informed as more information is available. What is the detour route? Road users will be detoured from Broken Hill to Wilcannia on the Barrier Highway, onto the Cobb Highway to Hay, then Sturt Highway to Wentworth while the Silver City Highway is closed at this location. The distance between Broken Hill and Wentworth on the Silver City Highway is 267 kilometres. The detour between the towns via the detour route is 963 kilometres, adding an extra nine hours to journeys. Motorists are advised to take this into consideration when planning their journey. Why has this route been chosen? Despite adding nine hours to journeys, the detour is the most suitable route around Bunnerungee Bridge when one considers road condition and communications. It is also a nominated heavy vehicle route, accessible by all vehicle types. There are two local road light vehicle alternatives around the bridge – via Anabranch and Pooncarie, adding 2½ and almost 4 hours respectively. -

New South Wales Class 1 Agricultural Vehicles (Notice) 2015 (No
Commonwealth of Australia Gazette Published by the Commonwealth of Australia GOVERNMENT NOTICES HEAVY VEHICLE NATIONAL LAW New South Wales Class 1 Agricultural Vehicles (Notice) 2015 (No. 1) This notice revokes the Class 1 Agricultural Vehicles Notice 2014 published in the New South Wales Government Gazette No. 15 of 7 February 2014, at page 426 to 459 and replaces it with Schedule 1. 1 Purpose (1) The purpose of this notice is to exempt the stated categories of class 1 heavy vehicles from the prescribed mass and dimension requirements specified in the notice subject to the conditions specified in the notice. 2 Authorising Provision(s) (1) This notice is made under Section 117, and Section 23 of Schedule 1, of the Heavy Vehicle National Law as in force in each participating jurisdiction. 3 Title (1) This notice may be cited as the New South Wales Class 1 Agricultural Vehicles (Notice) 2015 (No. 1) 4 Period of operation (1) This notice commences on the date of its publication in the Commonwealth Gazette and is in force for a period of five years from and including the date of commencement. 5 Definitions and interpretation (1) In this Instrument— (a) any reference to a provision of, or term used in, the former legislation, is to be taken to be a reference to the corresponding provision of, or nearest equivalent term used in, the Heavy Vehicle National Law; and (b) former legislation, means the Road Transport (Mass, Loading and Access) Regulation 2005 (NSW) and the Road Transport (Vehicle and Driver Management) Act 2005 (NSW); and (c) National Regulation means the Heavy Vehicle (Mass, Dimension and Loading) National Regulation. -
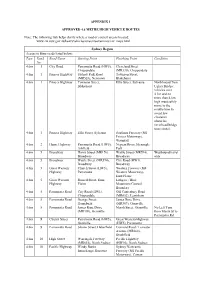
APPENDIX 1 APPROVED 4.6 METRE HIGH VEHICLE ROUTES Note: The
APPENDIX 1 APPROVED 4.6 METRE HIGH VEHICLE ROUTES Note: The following link helps clarify where a road or council area is located: www.rta.nsw.gov.au/heavyvehicles/oversizeovermass/rav_maps.html Sydney Region Access to State roads listed below: Type Road Road Name Starting Point Finishing Point Condition No 4.6m 1 City Road Parramatta Road (HW5), Cleveland Street Chippendale (MR330), Chippendale 4.6m 1 Princes Highway Sydney Park Road Townson Street, (MR528), Newtown Blakehurst 4.6m 1 Princes Highway Townson Street, Ellis Street, Sylvania Northbound Tom Blakehurst Ugly's Bridge: vehicles over 4.3m and no more than 4.6m high must safely move to the middle lane to avoid low clearance obstacles (overhead bridge truss struts). 4.6m 1 Princes Highway Ellis Street, Sylvania Southern Freeway (M1 Princes Motorway), Waterfall 4.6m 2 Hume Highway Parramatta Road (HW5), Nepean River, Menangle Ashfield Park 4.6m 5 Broadway Harris Street (MR170), Wattle Street (MR594), Westbound travel Broadway Broadway only 4.6m 5 Broadway Wattle Street (MR594), City Road (HW1), Broadway Broadway 4.6m 5 Great Western Church Street (HW5), Western Freeway (M4 Highway Parramatta Western Motorway), Emu Plains 4.6m 5 Great Western Russell Street, Emu Lithgow / Blue Highway Plains Mountains Council Boundary 4.6m 5 Parramatta Road City Road (HW1), Old Canterbury Road Chippendale (MR652), Lewisham 4.6m 5 Parramatta Road George Street, James Ruse Drive Homebush (MR309), Granville 4.6m 5 Parramatta Road James Ruse Drive Marsh Street, Granville No Left Turn (MR309), Granville -

On Road Rides Around Mildura
ON ROAD RIDES AROUND MILDURA Contents The Rides 1. Riverfront Circuit : Apex Park , Chaffey Bridge, 19 km 2. The Cemetery Circuit, 15-18.5 km 3. The “Spiders Web”: Red Cliffs and return, 38 km 4. The South West Loop : Benetook Rd, Werrimull Rd, 59 km 5 Nangiloc Tavern, 94 km 6. Cardross Loop, 35 km 7. Bridge to Bridge: Chaffey Bridge to Abbotsford Bridge, 46 km 8. The Arumpo/Mungo Road, 55 km 9. Werrimull Hotel and Lake Cullulleraine, 130 km 10. Lake Hawthorn, Golf Course and Airport, 34 Kms 11. Sunraysia “Hills”: Pump Hill , Merbein, 38 km 12. Trentham Estate Winery, 29 Km 13. The Chaffey History Trail, 10 km with extra options Attraction Finder Abbotsford Historical span lift bridge : ride 7 Apex Park Caravan park (and café) : ride 1 Gol Gol Hotel : ride 12 Historical Mildura and environs : ride 13 Inland Botanical Gardens : ride 7 Kings Billabong Lookout : ride 3 Mildura Homestead : ride 1, ride 13 Merbein Historical sites : ride 13 Nangiloc Tavern : ride 5 Psyche Pumps : ride 13 River Road Pottery /Café : ride 7 Trentham Cliffs Winery : ride 12 Visitor Centre : ride 13 Werrimull Hotel : ride 9 Woodsies Rock Shop : ride 3 About this ride guide There are many excellent rides on relatively quiet sealed roads around the Mildura district, this guide describes just thirteen of them. Although they are described as separate rides, quite a few, particularly those travelling further out of Mildura incorporate elements of other described rides. Rides can also be linked to increase distance. All rides, with the exception of the Chaffey Trail ride, depart from the City Centre at Langtree Mall and Ninth Sts. -

New South Wales Class 1 Special Purpose Vehicle and Combination Operator’S Guide
New South Wales Class 1 Special Purpose Vehicle and Combination Operator’s Guide 27 October 2020 New South Wales Class 1 Special Purpose Vehicle and Combination Operator’s Guide Contents Purpose .......................................................................................................................................................................................................... 3 Approved Routes and Travel Restrictions ..................................................................................................................................................... 3 1. Part 1 NSW Urban Zone ....................................................................................................................................................................... 3 1.1. Travel Restrictions in the NSW Urban Zone ................................................................................................................................... 3 1.1.1. Clearway and transit lane travel ............................................................................................................................................... 3 1.1.2. Peak hour travel ........................................................................................................................................................................ 4 1.1.3. Peak hour travel – Newcastle Outer Zone ................................................................................................................................ 4 1.1.4. Night travel .............................................................................................................................................................................. -

Wentworth Shire Council Local Strategic
––– Foreword Wentworth Shire Council has had the privilege of working with the Department of Planning, Industry & Environment Chief Planner’s office and Western Region staff to prepare this Local Strategic Planning Statement (LSPS). The purpose of the LSPS is to develop a vision for land use in the Shire for the next 20 years by recognising and understanding the current land use trends and anticipate those that may likely emerge. Our LSPS identifies three trends that have the ability to play a significant role in our future population and economic growth. These trends include changes to our agricultural industry, the demand and opportunities for new tourism enterprises and new residential development. Wentworth Shire Council is committed to achieving the goals of the Community Strategic Plan. We believe the LSPS will provide the mechanism to support and facilitate our economic growth, social cohesion and environmental sustainability for the next twenty years and beyond. Cr Melisa Hederics MAYOR WENTWORTH SHIRE Version Date Approved by Draft v1 13 November 2019 Michele Bos Draft v2 6 March 2020 Michele Bos 2 Acknowledgement of Country Wentworth Shire Council acknowledges the Traditional Owners of the land and pay respect to Elders past, present and emerging. Figure 1 - "Mungo - meeting place" by Craig Charles, Mutthi Mutthi This painting represents spirit and place – the coming together of three tribes, the Mutthi Mutthi, Paakantji, and Ngiyampaa. The centre of the spiral represents the middle of the Lake, the Walls of China and sacred sites. The many captions of gold leaf on the black swirls signify special ‘sites’ – uncovered and swept away by the winds. -

Historic Walking & Driving Tours
DARETON CALDER HWY CALDER SILVER CITY HWY TO CULLULLERAINE WERRIMULL Psyche Pumps MERINGUR 9 ADELAIDE The Coffee Palace STURT HWY Trace the story of how Mildura became an irrigated oasis in the midst of an arid land APEX PARK FLORA AVE DYAR AVE RIVERSIDE AVE ONTARIO AVE HistoricWALNUT Walking AVE & Mildura Wharf DrivingDEAKIN AVE Tours15TH ST/CALDER HWY (TO MELBOURNE) SAN MATEO AVE ETIWANDA AVE BENETOOK AVE COWRA AVE 11TH ST 14TH ST 5TH ST 19TH ST SANDILONG AVE 20TH ST KARADOC AVE Rio Vista KOORLONG KOORLONG AVE IRYMPLE AVE MORPUNG AVE GINQUAM AVE For more information please email [email protected] BELAR AVE For more history information please email [email protected] BELAR AVE DEWRY AVE May, 2015 CURETON AVE TO APEX PARK Drive 4 - Nichols Point Cemetery RIO VISTA NOWINGI PLACE & WATER PLAY Time, distance and map should be used as a guide only. WATER TOWER WATER Drive 5 - Red Cliffs U Murray Ave U The Chaffey Trail Reference Group in collaboration with the Mildura and District Historical Society with support from Merbein and Red Cliffs Historical Societies believe this to be an accurate document as at the time of printing 1 May 2015 Red Cliffs Streets: Rivers names run in alphabetical order east-west direction. Tree names run in alphabetical order, run north-south direction. MILDURA CENTRAL 73 SIXTEENTH STREET SEVENTEENTH STREET * This is a concept map, not to scale and to be used as a guide only for todays tour. Walks - Time and distance should be used as a guide only 1 - Historic Deakin Avenue Page 2 2 - Exploring East Mildura Page 10 3 - Mildura’s Riverfront history Page 12 4 - Rio Vista and surrounds Page 16 5 - The living heart of Mildura Page 19 Drives - Time and distance should be used as a guide only 1 - Mildura City Page 23 2 - Nichols Point Area Page 30 3 - Koorlong, Birdwoodton & Merbein Page 38 4 - Nichols Point Cemetery (Mildura) Page 49 5 - Red Cliffs Page 67 Foreword It is opportune to have the historic drives and walks back in print under the banner of the Chaffey Trail. -

New South Wales Government Gazette No. 26 of 28 June 2013
2989 Government Gazette OF THE STATE OF NEW SOUTH WALES Number 75 Monday, 24 June 2013 Published under authority by the Department of Premier and Cabinet SPECIAL SUPPLEMENT ROAD TRANSPORT (GENERAL) ACT 2005 ERRATUM IN New South Wales Government Gazette No. 25, dated 21st June 2013, at Folios 2549-2576 and Folios 2577-2604, a duplication of the fi rst Road Transport (General) Act 2005 notice has occurred in error. The fi rst notice on Folios 2519-2548 remains valid. 2990 SPECIAL SUPPLEMENT 24 June 2013 Road Transport (General) Act 2005 Class 1 Agricultural Vehicles (Amendment) Notice 2013 I, PETER WELLS, Director, Customer and Compliance Division, Roads and Maritime Services, pursuant to clause 12 of the Road Transport (Mass, Loading and Access) Regulation 2005 hereby amend the Class 1 Agricultural Vehicles Notice 2012 that was published on 27 July 2012 in the New South Wales Government Gazette No. 78 at pages 3428 to 3455 as set out in the Schedule of this Notice. The Class 1 Agricultural Vehicles (Amendment) Notice 2012 that was published on 2 November 2012 in the New South Wales Government Gazette No. 116 at pages 4526 to 4549 is hereby revoked. PETER WELLS Director, Customer and Compliance Roads and Maritime Services SCHEDULE The Class 1 Agricultural Vehicles Notice 2012 that was published on 27 July 2012 in the New South Wales Government Gazette No. 78 at pages 3428 to 3455 remains in force subject to the following amendments: [1] Delete the words “and Table 4” in clause 4.2.1 and clause 4.2.2 and in the Note that immediately follows Table 3.