1. Introduction
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Newsletter 123 May 2012
TheTheThe Irish Garden Plant Society Newsletter Number 11123123 May 2012 The Annual General Meeting 2012 The Annual General Meeting will be held on Sat 12th May 10.00 a.m for 10.30 a.m., at Hillsborough Courthouse, The Square, Hillsborough, BT26 6AG. As always, it will be followed by a series of garden visits on Saturday & Sunday and a meal on Saturday evening. The meal will be held at 8:00pm in La Mon Hotel & Country Club, 41 Gransha Road, Comber, BT23 5RF. See the January 2012 newsletter for details of the gardens to be visited. If you haven’t already booked contact Patrick Quigley, 24 Areema Drive, Dunmurry, Belfast, BT17 0QG. tel: +44 (0) 7801 299263 [email protected] for further information. A.G. M. Agenda 1. Apologies 2. Minutes of AGM 2011 3. Matters arising 4. Chairman’s report 5. Treasurer’s report 6. Election of Committee Members 7. Any other business Front cover : Solanum crispum ‘Glasnevin’. Photograph : Pearse Rowe In this issue 2 Editorial 3 Southern Climbers for Northern Walls by John Joe Costin 11 Worth a Read by Paddy Tobin 14 The Palm House, a review by Mary Bradshaw 16 Cheers to Chiltern Seeds and to the Sole Survivor of Seed Project 1997/98 by Michael Kelleher 17 Seed Distribution Report 2011 and 2012 by Stephen Butler 19 Regional Reports 28 Spring at Kilmacurragh by Seamus O’Brien 37 Propagation of Arbutus by Kevin Line 41 Tulipa ‘Molly Bloom’ - an new Tulip for 2012 1 Editorial Spring time and new plants are synonymous, and as Christopher Lloyd said in Garden Flowers from Seed, seed sowing is “one of life’s big thrills”. -
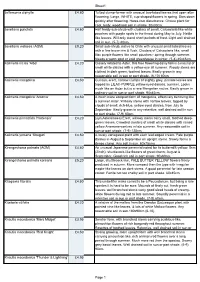
Sheet1 Jeffersonia Diphylla £4.80 Tufted Clump-Former with Unusual Two-Lobed Leaves That Open After Flowering
Sheet1 Jeffersonia diphylla £4.80 Tufted clump-former with unusual two-lobed leaves that open after flowering. Large, WHITE, cup-shaped flowers in spring. Dies down quickly after flowering. Hates root disturbance. Choice plant for cool, leafy, woodland soil in shade. 30x30cm. Jovellana punctata £4.60 Half hardy sub-shrub with clusters of small, Calceolaria like white pouches with purple spots in the throat during May to July. Nettle like leaves. Will only stand short periods of frost. Light well drained soil in sun. (5-7) 40cm. Jovellana violacea (AGM) £8.20 Small sub-shrub, native to Chile with unusual small lobed leaves with a fine brown rim & flush. Clusters of Calceolaria like, small lilac purple flowers like small pouches;- spring through summer. Needs a warm spot or cold greenhouse in winter. (5-8) 45x45cm. Kalimeris incisa 'Alba' £4.20 Closely related to Aster, this free flowering daisy forms a mound of small white daisies with a yellow eye all summer. Woody stems are clothed in dark green, toothed leaves. Easily grown in any reasonable soil in sun or part shade. (6-10) 60cm Kalimeris mongolica £4.50 Curious, erect, narrow clumps of slightly grey, pinnate leaves are topped by LILAC-PURPLE yellow-eyed daisies; summer. Looks much like an Aster but is a rare Mongolian native. Easily grown in ordinary soil in sun or part shade. 90x45cm. Kalimeris mongolica 'Antonia' £4.50 A much more compact form of mongolica, effectively behaving like a summer Aster. Willowy stems with narrow leaves, topped by clouds of small, rich-blue, yellow eyed daisies, from July to September. -

Show Schedules 2012 Ver Finale
119. 1 pan rock plant native to the Southern Hemisphere 120. 1 pan dwarf shurb THE SCOTTISH ROCK GARDEN CLUB 121. 1 pan rock plant raised from seed by the exhibitor. Date of sowing to be stated. Botanical notes permitted, AGS note 23(e) SECTION III Open to Amateur Members of AGS and SRGC who have not won an AGS Bronze Merit Medal or more than ten First Prizes at Shows run by either Society prior to 1st January 2011. Pan size not to exceed 19 cm outside diameter 130. 3 pans rock plants, distinct 131. 1 pan rock plant in flower 132. 1 pan Gentiana 133. 1 pan Cyclamen 134. 1 pan bulbous plant 135. 1 pan rock plant native to the Southern Hemisphere 136. 1 pan rock plant native to the Northern Hemisphere 137. 1 pan rock plant for foliage effect 138. 1 pan dwarf shrub or conifer 139. 1 pan rock plant. For exhibitors who have never won a first prize at an AGS or SRGC National show SHOW SCHEDULES 2012 DUNBLANE EARLY BULB DISPLAY 18th February* BLACKPOOL SHOW 17th March* STIRLING SHOW 24th March† New Location - Show this Year is in KINCARDINE NORTHUMBERLAND 40th ANNIVERSARY SHOW, HEXHAM 31st March EDINBURGH & THE LOTHIANS SHOW 14th April* PERTH SHOW 21st April HIGHLAND SHOW, NAIRN 28th April GLASGOW SHOW 5th May* ABERDEEN SHOW 19th May* GARDENING SCOTLAND (Joint Rock Only) 2nd June* LATE BULB DISPLAY, RBGE 8th September DISCUSSION WEEKEND, DUMFRIES 29th - 30th September NEWCASTLE SHOW 13th October* AGM 10th November† *Joint Rock Garden Plant Committee meetings 48 †Photographic/Art Competition SHOWS 2012 SHOW RULES 1. -

Temporal and Spatial Origin of Gesneriaceae in the New World Inferred from Plastid DNA Sequences
bs_bs_banner Botanical Journal of the Linnean Society, 2013, 171, 61–79. With 3 figures Temporal and spatial origin of Gesneriaceae in the New World inferred from plastid DNA sequences MATHIEU PERRET1*, ALAIN CHAUTEMS1, ANDRÉA ONOFRE DE ARAUJO2 and NICOLAS SALAMIN3,4 1Conservatoire et Jardin botaniques de la Ville de Genève, Ch. de l’Impératrice 1, CH-1292 Chambésy, Switzerland 2Centro de Ciências Naturais e Humanas, Universidade Federal do ABC, Rua Santa Adélia, 166, Bairro Bangu, Santo André, Brazil 3Department of Ecology and Evolution, University of Lausanne, CH-1015 Lausanne, Switzerland 4Swiss Institute of Bioinformatics, Quartier Sorge, CH-1015 Lausanne, Switzerland Received 15 December 2011; revised 3 July 2012; accepted for publication 18 August 2012 Gesneriaceae are represented in the New World (NW) by a major clade (c. 1000 species) currently recognized as subfamily Gesnerioideae. Radiation of this group occurred in all biomes of tropical America and was accompanied by extensive phenotypic and ecological diversification. Here we performed phylogenetic analyses using DNA sequences from three plastid loci to reconstruct the evolutionary history of Gesnerioideae and to investigate its relationship with other lineages of Gesneriaceae and Lamiales. Our molecular data confirm the inclusion of the South Pacific Coronanthereae and the Old World (OW) monotypic genus Titanotrichum in Gesnerioideae and the sister-group relationship of this subfamily to the rest of the OW Gesneriaceae. Calceolariaceae and the NW genera Peltanthera and Sanango appeared successively sister to Gesneriaceae, whereas Cubitanthus, which has been previously assigned to Gesneriaceae, is shown to be related to Linderniaceae. Based on molecular dating and biogeographical reconstruction analyses, we suggest that ancestors of Gesneriaceae originated in South America during the Late Cretaceous. -

Universidade Estadual Paulista Câmpus De
Campus de Botucatu UNESP - UNIVERSIDADE ESTADUAL PAULISTA CÂMPUS DE BOTUCATU INSTITUTO DE BIOCIÊN CIAS GENÔMICA ORGANELAR E EVOLUÇÃO DE GENLISEA E UTRICULARIA (LENTIBULARIACEAE) SAURA RODRIGUES DA SILVA Tese apresentada ao Instituto de Biociências, Câmpus de Botucatu, UNESP, para obtenção do título de Doutor em Ciências Biológicas (Botânica) BOTUCATU - SP - 2018 - Insti tuto de Biociências – Departamento de Botânica Distrito de Rubião Júnior s/n CEP 18618 - 000 Botucatu SP Brasil Tel 14 3811 6265/6053 fax 14 3815 3744 [email protected] Campus de Botucatu UNESP - UNIVERSIDADE ESTADUAL PAULISTA CÂMPUS DE BOTUCATU INSTITUTO DE BIOCIÊN CIAS GENÔMICA ORGANELAR E EVOLUÇÃO DE GENLISEA E UTRICULARIA (LENTIBULARIACEAE) SAUR A RODRIGUES DA SILVA PROF. DR. VITOR FERN ANDES OLIVEIRA DE MI RANDA ORIENTADOR PROF. DR. ALESSANDRO DE MEL L O VARANI Coorientador Tese apresentada ao Instituto de Biociências, Câmpus de Botucatu, UNESP, para obtenção do título de Doutor em Ciên cias Biológicas (Botânica) BOTUCATU - SP - 2018 - Instituto de Biociências – Departamento de Botânica Distrito de Rubião Júnior s/n CEP 18618 - 000 Botucatu SP Brasil Tel 14 3811 6265/6053 fax 14 3815 3744 [email protected] 2 Campus de Botucatu FICHA CATALOGRÁFIC A ELABORADA PELA SEÇÃO TÉCNICA DE AQUISIÇÃO E TRATAMENTO DA INFORMAÇÃO DIVISÃO TÉCNICA DE BIBLIOTECA E DOCUMENTAÇÃO - CAMPUS DE BOTUCATU - UNESP Silva, Saura Rodrigues. Genômica organelar e evolução d e Genlisea e Utricularia (Lentibulariaceae) / Saura Rodrigues da Silva. – 2018. Tese (doutorado) – Universidade Estadual Paulista, Instituto de Biociências de Botucatu, 2018. Orientador: Vitor Fernandes Oliveira de Miranda Co - orientador: Alessandro de Mello Varani Assunto CAPES: 1. Sistemática Vegetal CDD 581.1 Palavras - c have: Utricularia ; Genlisea ; genômica de organelas; ndhs; Evolução de organelas. -

Hidden in Plain Sight—A New Species of Lichen Strigula Oleistrata March 2020 (Strigulaceae) from New Zealand
TRILEPIDEA Newsletter of the New Zealand Plant Conservation Network NO. 196 Hidden in Plain sight—a new species of lichen Strigula oleistrata March 2020 (Strigulaceae) from New Zealand. Deadline for next issue: Marley Ford ([email protected]); Dan J. Blanchon ([email protected]), Friday 19 April 2020 School of Environmental & Animal Sciences, Unitec Institute of Technology, Auckland; SUBMIT AN ARTICLE Peter J. de Lange ([email protected]), School of Environmental & Animal TO THE NEWSLETTER Sciences, Unitec Institute of Technology, Auckland Contributions are welcome New Zealand has a surprising diversity of Strigula. 25 species of the genus are found to the newsletter at any here out of the c.70 species recognised; representing over a third of the known species time. The closing date for articles for each issue is (Galloway 2007, Lücking 2008, Hyde et al. 2013). Most of these species are foliicolous, approximately the 15th of meaning that they live on the surfaces of leaves. However, a few species do colonise each month. rocks and bark (Galloway 2007). Articles may be edited and used in the newsletter and/ Between 2016 and 2017 the senior author undertook a third year, level 7 School of or on the website news page. Environmental & Animal Sciences self-directed paper studying Strigula novae- The Network will publish zelandiae at the Unitec Institute of Technology Herbarium (UNITEC). Strigula novae- almost any article about zelandiae is a foliicolous species that is sometimes known as ‘silver paint lichen’, because plants and plant conservation with a particular focus on the when dead the thallus imparts a silvery patterning on the leaves it has colonised. -

Towards Resolving Lamiales Relationships
Schäferhoff et al. BMC Evolutionary Biology 2010, 10:352 http://www.biomedcentral.com/1471-2148/10/352 RESEARCH ARTICLE Open Access Towards resolving Lamiales relationships: insights from rapidly evolving chloroplast sequences Bastian Schäferhoff1*, Andreas Fleischmann2, Eberhard Fischer3, Dirk C Albach4, Thomas Borsch5, Günther Heubl2, Kai F Müller1 Abstract Background: In the large angiosperm order Lamiales, a diverse array of highly specialized life strategies such as carnivory, parasitism, epiphytism, and desiccation tolerance occur, and some lineages possess drastically accelerated DNA substitutional rates or miniaturized genomes. However, understanding the evolution of these phenomena in the order, and clarifying borders of and relationships among lamialean families, has been hindered by largely unresolved trees in the past. Results: Our analysis of the rapidly evolving trnK/matK, trnL-F and rps16 chloroplast regions enabled us to infer more precise phylogenetic hypotheses for the Lamiales. Relationships among the nine first-branching families in the Lamiales tree are now resolved with very strong support. Subsequent to Plocospermataceae, a clade consisting of Carlemanniaceae plus Oleaceae branches, followed by Tetrachondraceae and a newly inferred clade composed of Gesneriaceae plus Calceolariaceae, which is also supported by morphological characters. Plantaginaceae (incl. Gratioleae) and Scrophulariaceae are well separated in the backbone grade; Lamiaceae and Verbenaceae appear in distant clades, while the recently described Linderniaceae are confirmed to be monophyletic and in an isolated position. Conclusions: Confidence about deep nodes of the Lamiales tree is an important step towards understanding the evolutionary diversification of a major clade of flowering plants. The degree of resolution obtained here now provides a first opportunity to discuss the evolution of morphological and biochemical traits in Lamiales. -

Evolutionary History of Floral Key Innovations in Angiosperms Elisabeth Reyes
Evolutionary history of floral key innovations in angiosperms Elisabeth Reyes To cite this version: Elisabeth Reyes. Evolutionary history of floral key innovations in angiosperms. Botanics. Université Paris Saclay (COmUE), 2016. English. NNT : 2016SACLS489. tel-01443353 HAL Id: tel-01443353 https://tel.archives-ouvertes.fr/tel-01443353 Submitted on 23 Jan 2017 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. NNT : 2016SACLS489 THESE DE DOCTORAT DE L’UNIVERSITE PARIS-SACLAY, préparée à l’Université Paris-Sud ÉCOLE DOCTORALE N° 567 Sciences du Végétal : du Gène à l’Ecosystème Spécialité de Doctorat : Biologie Par Mme Elisabeth Reyes Evolutionary history of floral key innovations in angiosperms Thèse présentée et soutenue à Orsay, le 13 décembre 2016 : Composition du Jury : M. Ronse de Craene, Louis Directeur de recherche aux Jardins Rapporteur Botaniques Royaux d’Édimbourg M. Forest, Félix Directeur de recherche aux Jardins Rapporteur Botaniques Royaux de Kew Mme. Damerval, Catherine Directrice de recherche au Moulon Président du jury M. Lowry, Porter Curateur en chef aux Jardins Examinateur Botaniques du Missouri M. Haevermans, Thomas Maître de conférences au MNHN Examinateur Mme. Nadot, Sophie Professeur à l’Université Paris-Sud Directeur de thèse M. -

Flora of New Zealand and Conservation Strategies
Flora of New Zealand and Conservation Strategies November 2017 Hannah Streeter Merlin 714 1 Contents Acknowledgments 2 Daily Diary 3 Conclusions 47 Bibliography 49 Cost Breakdown 49 Acknowledgements I would like to thank several groups of people. The Merlin Trust for providing the generous financial support which meant this trip of a lifetime could happen. The staff at the Auckland Botanic gardens for welcoming me into their team and sharing their wisdom and expertise, for their great travel tips and warning about the dreaded sand flies. All of the people from the Department of conservation who have provided information and guidance when route planning and have answered all of my many questions graciously. Last but not least the people who took pictures of me hugging trees and pointing at cool plants, don’t worry most of them didn’t make the cut! 2 Daily Diary Tuesday 31st October So after what seems like almost a lifetime, two whole days in the air pretty much, I finally arrived in Auckland at 6:45 am. Then after a little wait to collect baggage and make it through security and baggage check I made it out of the airport. I would like to point out here that the baggage checks and customs declaration checks are much more thorough and in depth than they are in the UK. Where we would just collect our bag and walk straight through the nothing to declare line with no one checking anything, in New Zealand it’s a little different. Here there is a form to fill in and hand over when you get to the other side of bag pick up. -

Updated Angiosperm Family Tree for Analyzing Phylogenetic Diversity and Community Structure
Acta Botanica Brasilica - 31(2): 191-198. April-June 2017. doi: 10.1590/0102-33062016abb0306 Updated angiosperm family tree for analyzing phylogenetic diversity and community structure Markus Gastauer1,2* and João Augusto Alves Meira-Neto2 Received: August 19, 2016 Accepted: March 3, 2017 . ABSTRACT Th e computation of phylogenetic diversity and phylogenetic community structure demands an accurately calibrated, high-resolution phylogeny, which refl ects current knowledge regarding diversifi cation within the group of interest. Herein we present the angiosperm phylogeny R20160415.new, which is based on the topology proposed by the Angiosperm Phylogeny Group IV, a recently released compilation of angiosperm diversifi cation. R20160415.new is calibratable by diff erent sets of recently published estimates of mean node ages. Its application for the computation of phylogenetic diversity and/or phylogenetic community structure is straightforward and ensures the inclusion of up-to-date information in user specifi c applications, as long as users are familiar with the pitfalls of such hand- made supertrees. Keywords: angiosperm diversifi cation, APG IV, community tree calibration, megatrees, phylogenetic topology phylogeny comprising the entire taxonomic group under Introduction study (Gastauer & Meira-Neto 2013). Th e constant increase in knowledge about the phylogenetic The phylogenetic structure of a biological community relationships among taxa (e.g., Cox et al. 2014) requires regular determines whether species that coexist within a given revision of applied phylogenies in order to incorporate novel data community are more closely related than expected by chance, and is essential information for investigating and avoid out-dated information in analyses of phylogenetic community assembly rules (Kembel & Hubbell 2006; diversity and community structure. -

Palinotaxonomía De Scrophulariaceae Sensu Lato
Bol. Soc. Argent. Bot. 51 (2) 2016 M. M. Sosa y C. R. Salgado - Palinotaxonomía de ScrophulariaceaeISSN sensu 0373-580 lato X Bol. Soc. Argent. Bot. 51 (2): 299-321. 2016 VALOR TAXONÓMICO DEL POLEN EN SCROPHULARIACEAE SENSU LATO MARÍA DE LAS MERCEDES SOSA1,3 y CRISTINA R. SALGADO2,3 Resumen: Este trabajo es el resultado del estudio palinomorfológico de 32 especies pertenecientes a 19 géneros de Scrophulariaceae sensu lato. Además se realizó un registro exhaustivo de la información bibliográfica existente sobre la morfología polínica con el objetivo de analizar las relaciones entre las variables palinológicas y la clasificación sistemática actual. Los granos de polen analizados son: mónades, radiosimétricos e isopolares, de tamaño variable desde pequeños (11 µm) hasta medianos (51 µm), predominantemente esferoidales (P/E= 1), a veces suboblatos (P/E= 0,76), oblato-esferoidales (P/E= 0,95), prolatos (P/E=1,63) y perprolatos (P/E= 2,63). Inaperturados, 3-(4-5) colpados, 3-(4-5) colporados, espiraperturados, 2-8 sincolpados, 3-diploporados y pantoporados. La exina puede ser semitectada (retipilada, reticulada o estriada perforada) y tectada (psilada, foveolada, perforada, escábrida, rugulada, verrugosa, granulada, estriada o equinulada). Se realizó un análisis estadístico en base a una matriz de datos, compuesta por los caracteres palinológicos de 32 especies estudiadas sumado a la recopilación bibliográfica de 140 géneros y 218 especies pertenecientes a Scrophulariaceae sensu lato. El análisis de agrupamiento (UPGMA), confirmó que las diferencias palinomorfológicas apoyan la clasificación sistemática actual. Palabras clave: Morfología del polen, sistemática, euripolínico, Scrophulariaceae. Summary: Taxonomic value of pollen in Scrophulariaceae sensu lato. This paper is result of a study of pollen morphology of 32 species belonging to 19 genera of Scrophulariaceae sensu lato. -

The Systematic Revision of Chaetanthera Ruiz & Pav., and The
A systematic revision of Chaetanthera Ruiz & Pav., and the reinstatement of Oriastrum Poepp. & Endl. (Asteraceae: Mutisieae) Alison Margaret Robertson Davies München 2010 A systematic revision of Chaetanthera Ruiz & Pav., and the reinstatement of Oriastrum Poepp. & Endl. (Asteraceae: Mutisieae) Dissertation der Fakultät für Biologie der Ludwig-Maximilians-Universität München vorgelegt von Alison Margaret Robertson Davies München, den 03. November 2009 Erstgutachter: Prof. Dr. Jürke Grau Zweitgutachter: Prof. Dr. Günther Heubl Tag der mündlichen Prüfung: 28. April 2010 For Ric, Tim, Isabel & Nicolas Of all the floures in the meade, Thanne love I most those floures white and rede, Such as men callen daysyes. CHAUCER, ‘Legend of Good Women’, Prol. 43 (c. 1385) “…a traveller should be a botanist, for in all views plants form the chief embellishment.” DARWIN, ‘Darwin’s Journal of a Voyage round the World’, p. 599 (1896) Acknowledgements The successful completion of this work is due in great part to numerous people who have contributed both directly and indirectly. Thank you. Especial thanks goes to my husband Dr. Ric Davies who has provided unwavering support and encouragement throughout. I am deeply indebted to my supervisor, Jürke Grau, who made this research possible. Thank you for your support and guidance, and for your compassionate understanding of wider issues. The research for this study was funded by part-time employment on digital archiving projects coordinated via the Botanische Staatssammlung Munchen (INFOCOMP, 2000 – 2003; API- Projekt, 2005). Appreciative thanks go to the many friends and colleagues from both the Botanische Staatssammlung and the Botanical Institute who have provided scientific and social support over the years.