Fiskesamfunnet I Osensjøen, Trysil Og Åmot Kommuner, Hedmark
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Folketeljing 1900 for 0501 Lillehammer Kjøpstad
Folketeljing 1900 for 0501 Lillehammer kjøpstad Registreringssentral for historiske data, Universitetet i Tromsø 25.09.2014 Utskrift frå Digitalarkivet, Arkivverket si teneste for publisering av kjelder på internett: http://digitalarkivet.no Innhaldet er registrert av Registreringssentral for historiske data, Universitetet i Tromsø Innhald Løpande liste .................................. 9 Førenamnsregister ...................... 127 Etternamnsregister ...................... 155 Fødestadregister .......................... 183 Bustadregister ............................. 211 4 Folketeljingar i Noreg Det er halde folketeljingar i Noreg i 1769, 1801, 1815, 1825, 1835, 1845, 1855, 1865, 1870 (i nokre byar), 1875, 1885 (i byane), 1891, 1900, 1910, 1920, 1930, 1946, 1950, 1960, 1970, 1980, 1990 og 2001. Av teljingane før 1865 er berre ho frå 1801 nominativ, dvs. ho listar enkeltpersonar ved namn. Teljingane i 1769 og 1815-55 er numeriske, men med namnelistar i grunnlagsmateriale for nokre prestegjeld. Statistikklova i 1907 la sterke restriksjonar på bruken av nyare teljingar. Etter lov om offisiell statistikk og Statistisk Sentralbyrå (statistikklova) frå 1989 skal desse teljingane ikkje frigjevast før etter 100 år. 1910-teljinga blei difor frigjeven 1. desember 2010. Folketeljingane er avleverte til Arkivverket. Riksarkivet har originalane frå teljingane i 1769, 1801, 1815-1865, 1870, 1891, 1910, 1930, 1950, 1970 og 1980, mens statsarkiva har originalane til teljingane i 1875, 1885, 1900, 1920, 1946 og 1960 for sine distrikt. Folketeljinga 3. desember 1900 Ved kgl. res. 8. august 1900 blei det bestemt å halde ei "almindelig Folketælling" som skulle gje ei detaljert oversikt over befolkninga i Noreg natta mellom 2. og 3. desember 1900. På kvar bustad skulle alle personar til stades førast i teljingslista, med særskild markering ("mt") av dei som var mellombels til stades (på besøk osb.) på teljingstidspunktet. -

Norsk Natur 1-1968
~@~~~ NR. 1-4. ARGANG - MARS - 1968 , , >' REDAKT0R: RAGNAI<. FRISLID , r !l;~j-.; ~~&uM[ffi NORGES NATURVERNFORBUNDS TIDSSKRIFT ~. ,~ ~ Vellykt naturvernfremst0t i Tr0ndelag S0r-Tf!2lndelag Naturvern har i februar og mars i ar gjort et meget vellykket frem sl0t for a vekke interesse for naturverntanken, og for a verve medlemmer. Del 0ye blikkelige resultat av «Trondheims-aksjonen» var en 0kning av medlemstallet pa mer 500. Men det er klart at aksjonen har hatt langt videre virkninger, som ikke lar seg male i dag. Tusener pa tusener av s0rtr0ndere er blitt opmerksom pa hva naturvern er. Og det vii bety noe pa lang sikt. I forbindelse rned aksjonen sendte Tr0ndelag Naturvern ut sin vakre brosjyre «Moderne naturvern» av professor Ulf Hafsten. Aile nye rnedlemmer fikk dessuten forbundets brosjyre «Norsk naturvern i dap>. Under aksjonen ble det holdt foredrag i byen og distriktene, alle byens bokhandlere og Folkebiblioteket arrangerte spe sielle utstillinger over naturlitteratur, og plakater og avisannonsering ble tatt i bruk. Aksjonen be fulgt opp av avisene og av radioens lokalsendi.ng, og studenter ved to h0yskoler, speidere, medlemmer av Trondheim Feltbiologiske Ungdomsforening og andre deltok aktiv( i rnedlemsve.rving og annet arbeid. Vi kan ikke her ga i detalj nar det gjelder gjennornf0ringen av aksjonen. Men S0r-Tr0ndelag Naturvern vii utarbeide en rapport som vi regner med vii ve kke interesse i andre lokalforeninger. Vi haper at flest rnulig av de 0vrige kretsforen ingene vii f01ge S0r-Tr0ndelags eksempel, og bygge pa de erfaringene «Trond heims-aksjonen» har gitt. Morgendis over s01vbleke pytter - en stille bukt. NORGES NORSK NATUR A V INNHOLDET NATURVERNFORBUND Utkommer 4 ganger arlig Inventeringsvirksomhet Sianke traneben vader mellom vate siv mens 2 Akersgt. -

Sykehuset Innlandet Tynset English Title Inpatients’ Experiences with Somatic Hospitals in 2015
Utgitt av Folkehelseinstituttet Seksjon for brukeropplevd kvalitet i Kunnskapssenteret Tittel Pasienterfaringer med norske sykehus i 2015. Sykehuset Innlandet Tynset English title Inpatients’ experiences with somatic hospitals in 2015. Sykehuset Innlandet Tynset Ansvarlig Camilla Stoltenberg, direktør Forfattere Olaf Holmboe, prosjektleder, forsker, Folkehelsinstituttet Øyvind Andresen Bjertnæs, forskningsleder, Folkehelseinstituttet ISSN 1890-1565 Rapport 2016: 224 Publikasjonstype PasOpp-rapport Antall sider 18 (35 inklusiv vedlegg) Oppdragsgiver Helsedirektoratet Forsidefoto Colourbox Emneord(MeSH) Inpatients; patient satisfaction; health care surveys; questionnaires; hospitals; Norway Sitering Holmboe O, Bjertnæs OA. Pasienterfaringer med norske sykehus i 2015. Sykehuset Innlandet Tynset. [Inpatients’ experiences with somatic hospitals in 2015. Sykehuset Innlandet Tynset] PasOpp-rapport 2016: 224. Oslo: Folkehelseinstituttet, 2016. 2 Tidsserie for indikatorene (2011-2015) Innhold 3 Innhold Forord Nasjonalt kunnskapssenter for helsetjenesten har siden 2011 gjennomført årlige un- dersøkelser om pasienters erfaringer med døgnopphold på norske somatiske sykehus. Fra årsskiftet 2015-2016 ble Kunnskapssenteret innlemmet i Folkehelseinstituttet. Denne rapporten viser resultater fra en undersøkelse om pasienters erfaringer med sy- kehusopphold høsten 2015. Formålet er å formidle systematisk informasjon som kan brukes i kvalitetsforbedring og virksomhetsstyring. I store trekk har vi fulgt metoden som ble brukt i de tidligere undersøkelsene -

Lasting Legacies
Tre Lag Stevne Clarion Hotel South Saint Paul, MN August 3-6, 2016 .#56+0).')#%+'5 6*'(7674'1(1742#56 Spotlights on Norwegian-Americans who have contributed to architecture, engineering, institutions, art, science or education in the Americas A gathering of descendants and friends of the Trøndelag, Gudbrandsdal and northern Hedmark regions of Norway Program Schedule Velkommen til Stevne 2016! Welcome to the Tre Lag Stevne in South Saint Paul, Minnesota. We were last in the Twin Cities area in 2009 in this same location. In a metropolitan area of this size it is not as easy to see the results of the Norwegian immigration as in smaller towns and rural communities. But the evidence is there if you look for it. This year’s speakers will tell the story of the Norwegians who contributed to the richness of American culture through literature, art, architecture, politics, medicine and science. You may recognize a few of their names, but many are unsung heroes who quietly added strands to the fabric of America and the world. We hope to astonish you with the diversity of their talents. Our tour will take us to the first Norwegian church in America, which was moved from Muskego, Wisconsin to the grounds of Luther Seminary,. We’ll stop at Mindekirken, established in 1922 with the mission of retaining Norwegian heritage. It continues that mission today. We will also visit Norway House, the newest organization to promote Norwegian connectedness. Enjoy the program, make new friends, reconnect with old friends, and continue to learn about our shared heritage. -

Awaiting Breivik's Verdict
(Periodicals postage paid in Seattle, WA) TIME-DATED MATERIAL — DO NOT DELAY Taste of Norway Roots & Connections Sound Spirits Barneblad: Aquavit: Gode venner er engler som løfter Fun with balloons oss opp igjen når våre vinger har That’s the spirit! glemt hvordan det er å fly. for kids of all ages Read more on page 8 – Nils Bredesen Read more on page 10 Norwegian American Weekly Vol. 123 No. 26 June 29, 2012 Established May 17, 1889 • Formerly Western Viking and Nordisk Tidende $1.50 per copy Norway.com News Find more at www.norway.com Awaiting Breivik’s verdict News The recent Oil Fund move to July 22 trial ends buy property based from tax haven Luxembourg is under fire after 10 weeks from Norwegian-born French corruption hunter Eva Joly. “I of emotionally cannot understand how it’s pos- charged sible to set up a branch of the Oil Fund in a tax haven. It should testimonies be prohibited,” said a shocked Joly to Klassekampen. NOK STAFF COMPILATION 6.6 billion (almost USD 1.1 Norwegian American Weekly billion) was invested in compa- nies registered in tax havens the “The funeral gave me moti- Cayman Islands, Jersey, Guern- sey, and Luxembourg last year. vation. The multicultural Norway Answering Eva Joly’s censure, I’m so proud of. It showed that he Labor (Ap) Deputy Minister of failed. And we must be proud of it. Finance Hilde Singsaas stated Bano did not die in vain. She died there is nothing unusual about for the multicultural Norway.” establishing a property fund in Lara Rashid was one of the Luxembourg when investing in five witnesses to testify on the last more than one country. -

For Vinger Og Austmarka - Nr 5 - 2018 - 65
FOR VINGER OG AUSTMARKA - NR 5 - 2018 - 65. ÅRGANG LEDER «RAUSHET» OG VENTETID Høsten er over oss for fullt. Vi er inne i en tid med mange perspektiver. å akseptere at vi er ulike, at Naturen gjør sitt for å berike oss raust med sin fargeprakt. Dagene blir vi ikke er perfekte, og gi rom for kortere og mørket preger ettermiddagene. Vi går inn i en ventetid. egne og andres ulikheter. Tenk å få være et medmenneske Adventstiden kommer med alt den inneholder og jeg tenker at det er inn i denne ventetiden vi har foran oss. Få oppleve at andre gleder seg så viktig å ta vare på tiden vi har og de vi har rundt oss. I høst var det over ditt smil og dine besøk. Se naturen forandre karakter og kunne markering av verdensdagen for psykisk helse. Og tema var RAUSHET. glede seg over lys som tennes i høstmørket. Så viktig dag og så fint et tema. Vi har alle en psykisk helse og raushet vil styrke den på mange måter. Ordet «raushet», DET kan vi ta med oss Det er gjerne mange ting som skjer utover høsten og vinteren inn i inn i de mørke høst og vinterkvelder og inn i denne ventetiden. adventstiden. Det skal handles, bakes og førjulskonserter kommer på løpende bånd. Tilbudene er mange. Der er fort gjort å gå seg litt vill Hva er det å være raus? Hvordan kan vi i møte med oss selv og med i mangfoldet som er. Vi er et rikt samfunn. Vi kan dele med oss av andre mennesker vise raushet? Det behøver ikke koste noen ting. -
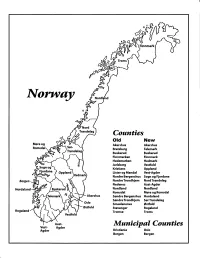
Norway Maps.Pdf
Finnmark lVorwny Trondelag Counties old New Akershus Akershus Bratsberg Telemark Buskerud Buskerud Finnmarken Finnmark Hedemarken Hedmark Jarlsberg Vestfold Kristians Oppland Oppland Lister og Mandal Vest-Agder Nordre Bergenshus Sogn og Fjordane NordreTrondhjem NordTrondelag Nedenes Aust-Agder Nordland Nordland Romsdal Mgre og Romsdal Akershus Sgndre Bergenshus Hordaland SsndreTrondhjem SorTrondelag Oslo Smaalenenes Ostfold Ostfold Stavanger Rogaland Rogaland Tromso Troms Vestfold Aust- Municipal Counties Vest- Agder Agder Kristiania Oslo Bergen Bergen A Feiring ((r Hurdal /\Langset /, \ Alc,ersltus Eidsvoll og Oslo Bjorke \ \\ r- -// Nannestad Heni ,Gi'erdrum Lilliestrom {", {udenes\ ,/\ Aurpkog )Y' ,\ I :' 'lv- '/t:ri \r*r/ t *) I ,I odfltisard l,t Enebakk Nordbv { Frog ) L-[--h il 6- As xrarctaa bak I { ':-\ I Vestby Hvitsten 'ca{a", 'l 4 ,- Holen :\saner Aust-Agder Valle 6rrl-1\ r--- Hylestad l- Austad 7/ Sandes - ,t'r ,'-' aa Gjovdal -.\. '\.-- ! Tovdal ,V-u-/ Vegarshei I *r""i'9^ _t Amli Risor -Ytre ,/ Ssndel Holt vtdestran \ -'ar^/Froland lveland ffi Bergen E- o;l'.t r 'aa*rrra- I t T ]***,,.\ I BYFJORDEN srl ffitt\ --- I 9r Mulen €'r A I t \ t Krohnengen Nordnest Fjellet \ XfC KORSKIRKEN t Nostet "r. I igvono i Leitet I Dokken DOMKIRKEN Dar;sird\ W \ - cyu8npris Lappen LAKSEVAG 'I Uran ,t' \ r-r -,4egry,*T-* \ ilJ]' *.,, Legdene ,rrf\t llruoAs \ o Kirstianborg ,'t? FYLLINGSDALEN {lil};h;h';ltft t)\l/ I t ,a o ff ui Mannasverkl , I t I t /_l-, Fjosanger I ,r-tJ 1r,7" N.fl.nd I r\a ,, , i, I, ,- Buslr,rrud I I N-(f i t\torbo \) l,/ Nes l-t' I J Viker -- l^ -- ---{a - tc')rt"- i Vtre Adal -o-r Uvdal ) Hgnefoss Y':TTS Tryistr-and Sigdal Veggli oJ Rollag ,y Lvnqdal J .--l/Tranbv *\, Frogn6r.tr Flesberg ; \. -

God Sommer! LEDER
FOR VINGER OG AUSTMARKA - NR 3 - 2019 - 66. ÅRGANG God sommer! LEDER BRY, BRYR, BRYSOM «Bry» er et fantastisk ord etter min hvor de trenger hjelp igjen. Men - det målestokk. Det er et relasjonelt ord. Det er også viktig å huske, at i de fleste er enten godt eller dårlig, for deg eller perioder er hjelp gjensidig, det kan meg. - Takk at du bryr deg / ikke bry deg, være en dobbelt glede å gi, både for - ikke plag meg. den som får og for den som gir – og neste gang kan det være omvendt – at Bryr du deg om å bruke blinklys? For å mottakeren gir og giveren får. Og HUSK fortelle folk hvor du har tenkt deg? – det du har gitt, kan ingen ta fra deg. Du valg, både kirkevalg og kommune- og Eller tenker du, at det trenger ingen er en gave til din samtid – la deg være fylkestingsvalg. Husk å bry deg – med andre bry seg med? Bryr du deg, om at en god gave til dem. Da kan du mest din stemme, - ditt engasjement for vei- du har mye eller lite – i forhold til hva sannsynlig høste gode relasjoner, når du valget, i både kommune, fylke og kirke. eller hvem? Bryr du deg om å være noe ønsker det. for andre? Eller å la andre være noe for Noen tror at de vet – andre vet at de deg? Er du til bry for andre? Eller er I Forkynneren kapittel 3 vers 1 står det tror. En overskrift i «Vårt land» andre til bry for deg? Ingen, kan være alt (I det gamle testamente i Bibelen): for alle – men alle kan være noe for noen. -

Årsrapport 2017 1 Yyyy Årsrapport 2017 HOVEDTALL 2017
Årsrapport 2017 1 yyyy Årsrapport 2017 HOVEDTALL 2017 Glommen Skog SA er et andelslag eid av 3 546 andelseiere med hovedkontor i Elverum. Selskapet ble etablert i 1903. MORSELSKAPET 2016 2017 Virkesomsetning Totalt tømmerkjøp i 1000 m3 1 683 1 723 Herav tømmerkjøp fra andelseiere i 1000 m3 1 221 1 277 Tømmeromsetning i mill. kroner 656 744 Leverandører antall 2 136 2 008 Gjennomsnittsleveranse pr leverandør i m3 784 858 Tjenester Adm. av skogsdrifter i 1000 m3 1 472 1 517 Omsetning av skogsdrift og skogkultur i mill. kr 265 270 Andelseiere Andelseiere antall 3 578 3 546 Skogeierområder antall 7 7 Andelseiernes produktive skogareal i 1000 daa 6 729 6 895 Ansatte Antall 72 71 Økonomi Driftsinntekter, mill. kroner 965 1 058 Driftsresultat, mill. kroner 3,8 3,7 Finansresultat, mill. kroner 21,8 29,1 Ordinært resultat før skatt, mill. kroner 25,6 32,8 Egenkapitalandel, % 89 86 Visjon Med kjærlighet til naturen og mennesket i sentrum skal vi sammen skape større verdier. Virksomhetsidé Vi er andelseiernes eget redskap for å utvikle størst mulig arealnytte. Formål Glommen Skog SAs formål er å bidra til best mulig avkastning på andels- eiernes eiendommer ved å; • tilby best mulig pris og avsetning for andelseiernes tømmer og et godt tilbud av konkurransedyktige tjenester, • investere i en rasjonell og konkurransedyktig skogindustri, • fremme vilkår som ivaretar og utvikler andelseiernes eiendoms- og forvaltningsrett, • la andelseierne ta del i den verdiutvikling som Glommen Skog SAs kapital får. Forsidebilde: Eyvind Wang Øvrige bilder: Tore Holaker og Glommens fotoarkiv. yyyy 2 3 yyyy Årsrapport 2017 TØMMEROMSETNING Med tømmeromsetning mener vi salg av tømmer, hogst ende for produsenter av grafisk trykkpapir, mens segmen- og transport. -

Menighetsbladet Sør-Odal Utgave 3 2019
Sør-Odal Menighetsblad 1 Høst 2019 Menighetsblad for Sør-Odal Årg. 80 / Nr. 3 Høst 2019 • Valgresultater Se side 3 • Skolestartgudstjeneste Se side 6 • Årets kirkemarsj Se side 12 Sør-Odal Menighetsblad 2 Høst 2019 Sør-Odal Menighetsblad 1 SØR-ODAL SØR-ODAL Høst 2019 SOKN KIRKEKONTOR Menighetsblad for Sør-Odal Årg. 80 / Nr. 3 Høst 2019 Adresse: Øgardsvegen 2, STRØM SOKN 2100 Skarnes Kirketjener: Bjørn E. Myhrer Tlf. 6297 8820 Faks: 6296 8088 Mobil: 900 23 986 Strøm menighetsråd Telefontid mandag-torsdag 9-15 Leder: Edel Lilleseth (tirsdag fra 12.00) • Valgresultater • Skolestartgudstjeneste • Årets kirkemarsj Se side 3 Se side 6 ULLERN SOKN Telefoner / e-poster Se side 12 Kirketjener: Bjarne Flåseth Kirkekontoret: 6297 8820 Mobil: 481 45 803 SØR-ODAL Ullern menighetsråd Assisterende kirkeverge: MENIGHETSBLAD Leder: Geir Kristoffersen Line Hemma 415 08 666 [email protected] Adresse: Kirkekontoret Øgardsvegen 2 OPPSTAD SOKN Kirkeverge: 2100 Skarnes Oppstad menighetsråd Asgeir Sektnan 958 18 858 Leder: Randi Bratt [email protected] E-post: [email protected] Sokneprester i Strøm, Ullern og Telefon: 6297 8820 SKARNES MENIGHETSHUS Oppstad: (Kirkekontoret) Anders Helset Eriksen Leder: Edel Lilleseth, Grendeveien, Mobil: 409 19 014 [email protected] Redaksjonskomité: 2100 Skarnes Kirkeverge Asgeir Sektnan (ansvarlig Tlf: 6296 1734 Jorunn Strand redaktør), sokneprest Anders Helset For leie av lokale: Mobil: 409 19 015 Eriksen, sokneprest Jorunn Strand, [email protected] Line Hemma, Kristin Lydia Nesse, Pedell: Leif Tafjord Elisabeth Vestgård. Strandvegen 9, Kantor 2100 Skarnes Kristin Lydia Nesse Konto for menighetsbladet: Mobil: 994 62 431 Mobil: 489 59 665 Odal [email protected] Sparebank: 1870.44.71811 Organist: Opplag: 3 700 Ahmad Mirzayev 994 65 162 Trykk: Flisa Trykkeri AS [email protected] Kantor Svein Tormod Bekkhus 905 04 091 STOFF TIL MENIGHETSBLADET [email protected] Vi tar gjerne imot stoff til Menighetspedagog: menighetsbladet! Elisabeth Vestgård 901 51 272 For å få det med i neste nummer, [email protected] send en e-post til [email protected] @ før 28. -

Hunting in Northern Europe Until 1500 AD Old Traditions and Regional
Hunting in northern Europe until 1500 AD Old traditions and regional developments, continental sources and continental influences The 7th century’s royal follower’s grave at mid-east Swedish Rickeby (Uppland) – the deceased one with his horse, several dogs, several raptor birds, several birds which represent the typical prey of falconry plus food gifts (drawing Ulla Malmsten). ScHriftEn DES ArcHäOlOgiScHEn lAnDESmuSEumS Ergänzungsreihe Band 7 Herausgegeben vom Archäologischen landesmuseum und dem Zentrum für Baltische und Skandinavische Archäologie in der Stiftung Schleswig-Holsteinische landesmuseen Schloss gottorf durch claus von carnap-Bornheim Hunting in northern Europe until 1500 AD Old traditions and regional developments, continental sources and continental influences Edited by Oliver grimm und ulrich Schmölcke Papers presented at a workshop organized by the centre for Baltic and Scandinavian Archaeology (ZBSA) Schleswig, June 16th and 17th, 2011 Wachholtz front matter illustration: after l. Sjösvärd/m. Vretemark/H. gustavson, A Vendel warrior from Vallentuna. in: J. P. lamm/ H.-Å nordström (eds.), Vendel Period Studies. transactions of the Boat-grave symposium Stockholm 1981 (Stockholm 1983) 133–150, fig. 5. iSBn 978 3 529 01877 0 redaktion: isabel Sonnenschein Satz und Bildbearbeitung: Jürgen Schüller, matthias Bolte Bibliografischei nformation der Deutschen nationalbibliothek. Die Deutsche nationalbibliothek verzeichnet diese Publikation in der Deutschen nationalbibliografie; detaillierte bibliografische Daten sind imi nternet -

For Vinger Og Austmarka - Nr 5 - 2017 - 64
FOR VINGER OG AUSTMARKA - NR 5 - 2017 - 64. ÅRGANG LEDER JULENS MYSTERIUM Førjulstida er ei koselig tid. Det er en sansetid full av lukter, Juleevangeliet er Guds smaker og lyder. Lukten av nybakte kaker fyller hjemmet vårt varme hilsen til oss. Det - pepperkaker, smultringer - ja, alle syv slags kaker. Smaken av er de gode nyhetene om at saftige og syrlige klementiner og glovarm gløgg med rosiner og vi ikke er forlatt, men at Gud mandel biter i. Førjulstida er ei koselig tid. har kommet til oss. Jesus var Guds Sønn og Verdens Frelser. Men Førjulstida er også mørketid. I et forsøk på å trenge mørket han var også et menneske - som oss. Det er julens mysterium. bort, pynter vi hjemmet vårt med levende lys, adventsstaker Det er gudstjenester med budskap om framtid og håp og med og julestjerner i vinduet. Vi tenner mange lys, samtidig som glade juletoner fra orgelet i hele desember. Mange kommer til det blir mørkere og mørkere ute. Små og store samles rundt kirken i denne tiden, men det er plass til flere. Det er plass til julegrana når den tennes i grender rundt omkring. deg! Kom og bli med og ta del i Julens mysterium! Julegrana minner oss om et annet tre; ”Den brennende tornebusken”. Gjennom den brennende tornebusken åpenbarte Med ønske om en velsignet høytid Gud seg og talte om trøst og håp da det var som mørkest for folket. Gud åpenbarte seg og talte om framtid og håp. Turid Lindal Barne- og ungdomsprest Vinger og Det er ikke alle som vet hva ”God Jul” betyr.