Thyroid Hormone Insufficiency Alters the Expression of Psychiatric
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

WSAVA List of Essential Medicines for Cats and Dogs
The World Small Animal Veterinary Association (WSAVA) List of Essential Medicines for Cats and Dogs Version 1; January 20th, 2020 Members of the WSAVA Therapeutic Guidelines Group (TGG) Steagall PV, Pelligand L, Page SW, Bourgeois M, Weese S, Manigot G, Dublin D, Ferreira JP, Guardabassi L © 2020 WSAVA All Rights Reserved Contents Background ................................................................................................................................... 2 Definition ...................................................................................................................................... 2 Using the List of Essential Medicines ............................................................................................ 2 Criteria for selection of essential medicines ................................................................................. 3 Anaesthetic, analgesic, sedative and emergency drugs ............................................................... 4 Antimicrobial drugs ....................................................................................................................... 7 Antibacterial and antiprotozoal drugs ....................................................................................... 7 Systemic administration ........................................................................................................ 7 Topical administration ........................................................................................................... 9 Antifungal drugs ..................................................................................................................... -

IHS National Pharmacy & Therapeutics Committee National
IHS National Pharmacy & Therapeutics Committee National Core Formulary; Last Updated: 09/23/2021 **Note: Medications in GREY indicate removed items.** Generic Medication Name Pharmacological Category (up-to-date) Formulary Brief (if Notes / Similar NCF Active? available) Miscellaneous Medications Acetaminophen Analgesic, Miscellaneous Yes Albuterol nebulized solution Beta2 Agonist Yes Albuterol, metered dose inhaler Beta2 Agonist NPTC Meeting Update *Any product* Yes (MDI) (Nov 2017) Alendronate Bisphosphonate Derivative Osteoporosis (2016) Yes Allopurinol Antigout Agent; Xanthine Oxidase Inhibitor Gout (2016) Yes Alogliptin Antidiabetic Agent, Dipeptidyl Peptidase 4 (DPP-4) Inhibitor DPP-IV Inhibitors (2019) Yes Anastrozole Antineoplastic Agent, Aromatase Inhibitor Yes Aspirin Antiplatelet Agent; Nonsteroidal Anti-Inflammatory Drug; Salicylate Yes Azithromycin Antibiotic, Macrolide STIs - PART 1 (2021) Yes Calcium Electrolyte supplement *Any formulation* Yes Carbidopa-Levodopa (immediate Anti-Parkinson Agent; Decarboxylase Inhibitor-Dopamine Precursor Parkinson's Disease Yes release) (2019) Clindamycin, topical ===REMOVED from NCF=== (See Benzoyl Peroxide AND Removed January No Clindamycin, topical combination) 2020 Corticosteroid, intranasal Intranasal Corticosteroid *Any product* Yes Cyanocobalamin (Vitamin B12), Vitamin, Water Soluble Hematologic Supplements Yes oral (2016) Printed on 09/25/2021 Page 1 of 18 National Core Formulary; Last Updated: 09/23/2021 Generic Medication Name Pharmacological Category (up-to-date) Formulary Brief -

Drugs Affectin the Autonomic Nervous System
Fundamentals of Medical Pharmacology Paterson Public Schools Written by Néstor Collazo, Ph.D. Jonathan Hodges, M.D. Tatiana Mikhaelovsky, M.D. for Health and Related Professions (H.A.R.P.) Academy March 2007 Course Description This fourth year course is designed to give students in the Health and Related Professions (H.A.R.P.) Academy a general and coherent explanation of the science of pharmacology in terms of its basic concepts and principles. Students will learn the properties and interactions between chemical agents (drugs) and living organisms for the rational and safe use of drugs in the control, prevention, and therapy of human disease. The emphasis will be on the fundamental concepts as they apply to the actions of most prototype drugs. In order to exemplify important underlying principles, many of the agents in current use will be singled out for fuller discussion. The course will include the following topics: ¾ The History of Pharmacology ¾ Terminology Used in Pharmacology ¾ Drug Action on Living Organisms ¾ Principles of Pharmacokinetics ¾ Dose-Response Relationships ¾ Time-Response Relationships ¾ Human Variability: Factors that will modify effects of drugs on individuals ¾ Effects of Drugs Attributable to Varying Modes of Administration ¾ Drug Toxicity ¾ Pharmacologic Aspects of Drug Abuse and Drug Dependence Pre-requisites Students must have completed successfully the following courses: Biology, Chemistry, Anatomy and Physiology, Algebra I and II Credits: 5 credits Basic Principles of Drug Action Introduction to Pharmacology a. Basic Mechanisms of Drug Actions b. Dose-response relationships c. Drug absorption d. Biotransformation of Drugs e. Pharmacokinetics f. Factors Affecting Drug Distribution g. Drug Allergy and Pharmacogenetics h. -

Repurposing of Drugs for Triple Negative Breast Cancer: an Overview
Repurposing of drugs for triple negative breast cancer: an overview Andrea Spini1,2, Sandra Donnini3, Pan Pantziarka4, Sergio Crispino4,5 and Marina Ziche1 1Department of Medicine, Surgery and Neuroscience, University of Siena, Siena 53100, Italy 2Service de Pharmacologie Médicale, INSERM U1219, University of Bordeaux, Bordeaux 33000, France 3Department of Life Sciences, University of Siena, Siena 53100, Italy 4Anticancer Fund, Strombeek Bever 1853, Belgium 5ASSO, Siena, Italy Abstract Breast cancer (BC) is the most frequent cancer among women in the world and it remains a leading cause of cancer death in women globally. Among BCs, triple negative breast cancer (TNBC) is the most aggressive, and for its histochemical and molecular charac- teristics is also the one whose therapeutic opportunities are most limited. The REpur- posing Drugs in Oncology (ReDO) project investigates the potential use of off patent non-cancer drugs as sources of new cancer therapies. Repurposing of old non-cancer drugs, clinically approved, off patent and with known targets into oncological indications, Review offers potentially cheaper effective and safe drugs. In line with this project, this article describes a comprehensive overview of preclinical or clinical evidence of drugs included in the ReDO database and/or PubMed for repurposing as anticancer drugs into TNBC therapeutic treatments. Keywords: triple negative breast cancer, repositioning, non-cancer drug, preclinical studies, clinical studies Background Correspondence to: Marina Ziche Breast cancer (BC) is the most frequent cancer among women in the world. Triple nega- Email: [email protected] tive breast cancer (TNBC) is a type of BC that does not express oestrogen receptors, pro- 2020, :1071 gesterone receptors and epidermal growth factor receptors-2/Neu (HER2) and accounts ecancer 14 https://doi.org/10.3332/ecancer.2020.1071 for the 16% of BCs approximatively [1, 2]. -

Smoking and Hormones in Health and Endocrine Disorders
European Journal of Endocrinology (2005) 152 491–499 ISSN 0804-4643 REVIEW Smoking and hormones in health and endocrine disorders D Kapoor and T H Jones Centre for Diabetes and Endocrinology, Barnsley District General Hospital, Gawber Road, Barnsley S75 2EP, UK and Academic Unit of Endocrinology, Division of Genomic Medicine, University of Sheffield, Sheffield, UK (Correspondence should be addressed to T H Jones; Email: [email protected]) Abstract Smoking has multiple effects on hormone secretion, some of which are associated with important clinical implications. These effects are mainly mediated by the pharmacological action of nicotine and also by toxins such as thiocyanate. Smoking affects pituitary, thyroid, adrenal, testicular and ovarian function, calcium metabolism and the action of insulin. The major salient clinical effects are the increased risk and severity of Graves’ hyperthyroidism and opthalmopathy, osteoporosis and reduced fertility. Smoking also contributes to the development of insulin resistance and hence type 2 diabetes mellitus. An important concern is also the effect of smoking on the foetus and young children. Passive transfer of thiocyanate can cause disturbance of thyroid size and function. Furthermore, maternal smoking causes increased catecholamine production, which may contribute to under perfusion of the foetoplacental unit. European Journal of Endocrinology 152 491–499 Introduction and also the liability of blood platelets to adhere to each other and to the walls of blood vessels. Nicotine The health consequences of cigarette smoking and of also causes stimulation and sedation of the central ner- the use of other tobacco products are well known. vous system depending upon the dose. -

Advances in Hepatology
AdvAnces In HepAtology Current Developments in the Treatment of Hepatitis and Hepatobiliary Disease Hepatology section editor: eugene R. schiff, Md The Adaptive Response (Drug Tolerance) Helps to Prevent Drug-Induced Liver Injury James H. Lewis, MD Professor and Director of Hepatology Division of Gastroenterology Department of Medicine Georgetown University Hospital Washington, DC G&H How frequently do drugs cause agents associated with drug tolerance. Up to 5% of elevations in liver-associated enzymes or lead patients taking statins will develop ALT elevations, which to drug-induced liver injury? usually remain less than 3 times the upper limit of normal (ULN) and are not associated with any hepatic-related JHL Elevations in liver-associated enzymes (LAEs)—in symptoms. Nevertheless, owing to the labeling that particular, alanine aminotransferase (ALT) and aspartate accompanied statin approval, which mentioned a risk of aminotransferase—are seen fairly frequently in clinical hepatotoxicity and need for LAE monitoring, even such practice and can be related to any number of acute and low-level elevations continue to cause consternation in chronic causes, including viral hepatitis, fatty-liver disease, the clinical setting, and the drug is often stopped prior to alcohol, a variety of autoimmune and metabolic disorders, determining if tolerance will develop. and several drugs. When it comes to abnormal liver test Other drugs are associated with a prevalence of sub- results and drug-induced liver injury (DILI), it is important clinical hepatic enzyme elevations in the 10–20% range, for clinicians to draw a distinction between asymptomatic, including a number of antibiotics, such as erythromycin low-level elevations in LAEs and biochemical changes estolate, ketoconazole, and isoniazid. -
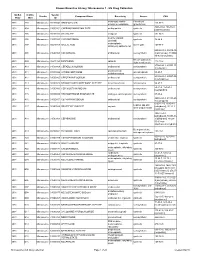
Known Bioactive Library: Microsource 1 - US Drug Collection
Known Bioactive Library: Microsource 1 - US Drug Collection ICCB-L ICCB-L Vendor Vendor Compound Name Bioactivity Source CAS Plate Well ID antifungal, inhibits Penicillium 2091 A03 Microsource 00200046 GRISEOFULVIN 126-07-8 mitosis in metaphase griseofulvum 3505-38-2, 486-16-8 2091 A04 Microsource 01500161 CARBINOXAMINE MALEATE antihistaminic synthetic [carbinoxamine] 2091 A05 Microsource 00200331 SALSALATE analgesic synthetic 552-94-3 muscle relaxant 2091 A06 Microsource 01500162 CARISOPRODOL synthetic 78-44-4 (skeletal) antineoplastic, 2091 A07 Microsource 00210369 GALLIC ACID insect galls 149-91-7 astringent, antibacterial 66592-87-8, 50370-12- 2091 A08 Microsource 01500163 CEFADROXIL antibacterial semisynthetic 2 [anhydrous], 119922- 89-9 [hemihydrate] Rheum palmatum, 2091 A09 Microsource 00211468 DANTHRON cathartic 117-10-2 Xyris semifuscata 27164-46-1, 25953-19- 2091 A10 Microsource 01500164 CEFAZOLIN SODIUM antibacterial semisynthetic 9 [cefazolin] glucocorticoid, 2091 A11 Microsource 00300024 HYDROCORTISONE adrenal glands 50-23-7 antiinflammatory 64485-93-4, 63527-52- 2091 A12 Microsource 01500165 CEFOTAXIME SODIUM antibacterial semisynthetic 6 [cefotaxime] 2091 A13 Microsource 00300029 DESOXYCORTICOSTERONE ACETATE mineralocorticoid adrenocortex 56-47-3 58-71-9, 153-61-7 2091 A14 Microsource 01500166 CEPHALOTHIN SODIUM antibacterial semisynthetic [cephalothin] 2091 A15 Microsource 00300034 TESTOSTERONE PROPIONATE androgen, antineoplastic semisynthetic 57-85-2 24356-60-3, 21593-23- 2091 A16 Microsource 01500167 CEPHAPIRIN SODIUM -

Bp501t Medchem -Unit
Bachelor of Pharmacy (B. Pharm) SEMESTER:5TH Subject: MEDICINAL CHEMISTRY-II CODE: BP501T UNIT:IV UNIT: IV 4. Drugs acting on Endocrine system 4.1. Introduction to steroids 4.1.1. Classification of Steroids 4.1.2. Nomenclature, 4.1.3. Stereochemistry 4.1.4. Metabolism of steroids 4.2. Sex hormones: 4.2.1. Testosterone, 4.2.2. Nandralone, 4.2.3. Progestrones, 4.2.4. Oestriol, 4.2.5. Oestradiol, 4.2.6. Oestrione, 4.2.7. Diethyl stilbestrol. 4.3. Drugs for erectile dysfunction 4.3.1. Sildenafil 4.3.2. Tadalafil 4.4. Oral contraceptives 4.4.1. Mifepristone 4.4.2. Norgestril 4.4.3. Levonorgestrol 4.5. Corticosteroids 4.5.1. Cortisone 4.5.2. Hydrocortisone 4.5.3. Prednisolone 4.5.4. Betamethasone 4.5.5. Dexamethasone 4.6. Thyroid and antithyroid drugs 4.6.1. L-Thyroxine 4.6.2. L-Thyronine 4.6.3. Propylthiouracil 4.6.4.Methimazole 4. Endocrine system The endocrine system helps to maintain internal homeostasis through the use of endogenous chemicals known as hormones. A hormone is typically regarded as a chemical messenger that is released into the bloodstream to exert an effect on target cells located some distance from the hormonal release site. The endocrine system is a series of glands that produce hormones which regulate respiration, metabolism, growth and development, tissue function, sexual function, reproduction etc. Endocrine glands are ductless glands of the endocrine system that secrete their products, hormones, directly into the blood. The major glands of the endocrine system include the pineal gland, pituitary gland, pancreas, ovaries, testes, thyroid gland, parathyroid gland, hypothalamus and adrenal glands (Fig. -

IHS National Pharmacy & Therapeutics Committee National
IHS National Pharmacy & Therapeutics Committee National Core Formulary; Last Updated: 09/23/2021 **Note: Medications in GREY indicate removed items.** Generic Pharmacological Category Formulary Brief (if Notes / Similar NCF Medications Active? Medication Name (up-to-date) available) Miscellaneous Acetaminophen Analgesic, Miscellaneous Yes Albuterol nebulized Beta2 Agonist Yes solution Alendronate Bisphosphonate Derivative Osteoporosis (2016) Yes Allopurinol Antigout Agent; Xanthine Oxidase Gout (2016) Yes Inhibitor Alogliptin Antidiabetic Agent, Dipeptidyl DPP-IV Inhibitors Yes Peptidase 4 (DPP-4) Inhibitor (2019) Amitriptyline Antidepressant, Tricyclic Antidepressants in Nortriptyline Yes Pain Mgmt (2014) Amlodipine Antihypertensive; Calcium Calcium Channel Diltiazem Yes Channel Blocker, Dihydropyridine Blockers (2014) Anastrozole Antineoplastic Agent, Aromatase Yes Inhibitor Apixaban Anticoagulant; Direct Oral Direct Oral Warfarin Yes Anticoagulant (DOAC); Factor Xa Anticoagulants (2017) Inhibitor Aspirin Antiplatelet Agent; Nonsteroidal Yes Anti-Inflammatory Drug; Salicylate Atenolol Antihypertensive; Beta-Blocker, Beta Blockers (2014) Metoprolol; Propranolol Yes Beta1 Selective Atomoxetine Norepinephrine Reuptake Inhibitor, ADHD (2020) Dextroamphetamine / Amphetamine (immediate release); Yes Selective Dextroamphetamine / Amphetamine (long-acting); Methylphenidate (immediate release); Methylphenidate (long-acting) Atorvastatin Antilipemic Agent, HMG-CoA Dyslipidemia Guideline Pravastatin; Rosuvastatin; Simvastatin Yes Reductase Inhibitor -

Systematic Assessment of Prescribed Medications and Short-Term
www.nature.com/scientificreports Corrected: Author Correction OPEN Systematic assessment of prescribed medications and short- term risk of myocardial infarction Received: 29 November 2018 Accepted: 14 May 2019 – a pharmacopeia-wide association Published online: 04 June 2019 study from Norway and Sweden Abhijit Sen1,2, Ioannis Vardaxis3, Bo Henry Lindqvist3, Ben Michael Brumpton4,5,6, Linn Beate Strand1, Inger Johanne Bakken 7, Lars Johan Vatten1, Pål Richard Romundstad1, Rickard Ljung8, Kenneth Jay Mukamal9 & Imre Janszky1,10 Wholesale, unbiased assessment of Scandinavian electronic health-care databases ofer a unique opportunity to reveal potentially important undiscovered drug side efects. We examined the short- term risk of acute myocardial infarction (AMI) associated with drugs prescribed in Norway or Sweden. We identifed 24,584 and 97,068 AMI patients via the patient- and the cause-of-death registers and linked to prescription databases in Norway (2004–2014) and Sweden (2005–2014), respectively. A case- crossover design was used to compare the drugs dispensed 1–7 days before the date of AMI diagnosis with 15–21 days’ time -window for all the drug individually while controlling the receipt of other drugs. A BOLASSO approach was used to select drugs that acutely either increase or decrease the apparent risk of AMI. We found 48 drugs to be associated with AMI in both countries. Some antithrombotics, antibiotics, opioid analgesics, adrenergics, proton-pump inhibitors, nitroglycerin, diazepam, metoclopramide, acetylcysteine were associated with higher risk for AMI; whereas angiotensin-II- antagonists, calcium-channel blockers, angiotensin-converting-enzyme inhibitors, serotonin-specifc reuptake inhibitors, allopurinol, mometasone, metformin, simvastatin, levothyroxine were inversely associated. -

Prescribing Information/Insert
Prescribing Information/Insert ELTROXINTM QUALITATIVE AND QUANTITATIVE COMPOSITION Each tablet contains 25, 50, 75, 88, 100, 112, 125, 137, 150, 175 µg of levothyroxine sodium Excipients Note for GSK operating companies: It is mandatory for country product information to include both the complete list of excipients for all locally marketed presentations, and any locally imposed excipient warning statements. PHARMACEUTICAL FORM Tablet CLINICAL INFORMATION Indications For the treatment of: • Hypothyroidism Supplier 2 For the treatment of: • benign, euthyroid goitre • co-therapy in the antithyroid treatment of hyperthyroidism, once euthyroid status has been achieved • thyroid suppression test Dosage and Administration The total daily dose should be taken in the morning on an empty stomach at least half an hour before breakfast. The tablets should be swallowed whole, without chewing, with some liquid. Due to lack of data, it is not appropriate to crush levothyroxine tablets. Levothyroxine tablets without a score-line must not be halved. Thyroid hormone therapy/ replacement The dosing information serves as a guideline. The individual daily dose should be determined by laboratory diagnostic tests and clinical examinations. If any residual thyroid function remains, a lower replacement dose may be sufficient. In elderly patients, patients with coronary heart disease and patients with severe or chronic hypothyroidism, thyroid hormone treatment must be initiated with particular caution, i.e. by selecting a low initial dose and increasing it slowly and at longer intervals, with frequent thyroid hormone monitoring. Experience has shown that a lower dose is also sufficient in patients with a low body weight and in patients with large goitres. A starting dose of 12.5 micrograms can be used, providing that available formulation allow for appropriate dose adaptation. -

Summary of Product Characteristics
SUMMARY OF PRODUCT CHARACTERISTICS 1. NAME OF THE MEDICINAL PRODUCT Levothyroxine Sodium {Aspen} 25 microgram tablets Levothyroxine Sodium {Aspen} 50 microgram tablets Levothyroxine Sodium {Aspen} 75 microgram tablets Levothyroxine Sodium {Aspen} 88 microgram tablets Levothyroxine Sodium {Aspen} 100 microgram tablets Levothyroxine Sodium {Aspen} 112 microgram tablets Levothyroxine Sodium {Aspen} 125 microgram tablets Levothyroxine Sodium {Aspen} 137 microgram tablets Levothyroxine Sodium {Aspen} 150 microgram tablets Levothyroxine Sodium {Aspen} 175 microgram tablets Levothyroxine Sodium {Aspen} 200 microgram tablets 2. QUALITATIVE AND QUANTITATIVE COMPOSITION Each tablet of Levothyroxine sodium {Aspen} 25 microgram contains 27.8 microgram of levothyroxine sodium pentahydrate equivalent to 25 microgram of levothyroxine sodium. Each tablet of Levothyroxine sodium {Aspen} 50 microgram contains 55.6 microgram of levothyroxine sodium pentahydrate equivalent to 50 microgram of levothyroxine sodium. Each tablet of Levothyroxine sodium {Aspen} 75 microgram contains 83.4 microgram of levothyroxine sodium pentahydrate equivalent to 75 microgram of levothyroxine sodium. Each tablet of Levothyroxine sodium {Aspen} 88 microgram contains 97.9 microgram of levothyroxine sodium pentahydrate equivalent to 88 microgram of levothyroxine sodium. Each tablet of Levothyroxine sodium {Aspen} 100 microgram contains 111.3 microgram of levothyroxine sodium pentahydrate equivalent to 100 microgram of levothyroxine sodium. Each tablet of Levothyroxine sodium {Aspen} 112 microgram contains 124.6 microgram of levothyroxine sodium pentahydrate equivalent to 112 microgram of levothyroxine sodium. Each tablet of Levothyroxine sodium {Aspen} 125 microgram contains 139.1 microgram of levothyroxine sodium pentahydrate equivalent to 125 microgram of levothyroxine sodium. Each tablet of Levothyroxine sodium {Aspen} 137 microgram contains 152.4 microgram of levothyroxine sodium pentahydrate equivalent to 137 microgram of levothyroxine sodium.