The Relationship Between Skull Morphology, Masticatory Muscle Force
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Questions on Human Anatomy
Standard Medical Text-books. ROBERTS’ PRACTICE OF MEDICINE. The Theory and Practice of Medicine. By Frederick T. Roberts, m.d. Third edi- tion. Octavo. Price, cloth, $6.00; leather, $7.00 Recommended at University of Pennsylvania. Long Island College Hospital, Yale and Harvard Colleges, Bishop’s College, Montreal; Uni- versity of Michigan, and over twenty other medical schools. MEIGS & PEPPER ON CHILDREN. A Practical Treatise on Diseases of Children. By J. Forsyth Meigs, m.d., and William Pepper, m.d. 7th edition. 8vo. Price, cloth, $6.00; leather, $7.00 Recommended at thirty-five of the principal medical colleges in the United States, including Bellevue Hospital, New York, University of Pennsylvania, and Long Island College Hospital. BIDDLE’S MATERIA MEDICA. Materia Medica, for the Use of Students and Physicians. By the late Prof. John B Biddle, m.d., Professor of Materia Medica in Jefferson Medical College, Phila- delphia. The Eighth edition. Octavo. Price, cloth, $4.00 Recommended in colleges in all parts of the UnitedStates. BYFORD ON WOMEN. The Diseases and Accidents Incident to Women. By Wm. H. Byford, m.d., Professor of Obstetrics and Diseases of Women and Children in the Chicago Medical College. Third edition, revised. 164 illus. Price, cloth, $5.00; leather, $6.00 “ Being particularly of use where questions of etiology and general treatment are concerned.”—American Journal of Obstetrics. CAZEAUX’S GREAT WORK ON OBSTETRICS. A practical Text-book on Midwifery. The most complete book now before the profession. Sixth edition, illus. Price, cloth, $6.00 ; leather, $7.00 Recommended at nearly fifty medical schools in the United States. -

The Development of the Human Maxilla, Vomer, and Paraseptal Cartilages
THE DEVELOPMENT OF THE HUMAN MAXILLA, VOMER, AND PARASEPTAL CARTILAGES. By Professor FAWCETT, M.D., University of Bristol. THE usually accepted descriptions of the development of the maxilla of man state that it arises by a number of separate centres-the number varying somewhat with the authority, likewise the situation of these centres. No description of the maxilla can be considered complete unless at the same time notice is taken of the manner of development of the premaxilla, which, of course, forms the anterior segment of the adult bone as usually interpreted. But the consideration of the development of the premaxilla may be left until that of the maxilla has been fully dealt with. Before breaking new ground, it may be well to state what are the usual statements with reference to the ossification of the maxilla. These statements are apparently for the most part based on work done by Callender, Toldt, Rambaud and Renault, and Bland Sutton, so far as concerns human anatomy. More recently Franklin Mall has given his views on the subject in the American Jouarnal of Anatomy, views based on observation of specimens treated by the "clearing" method of Schulze. So far as they go, these statements are in harmony with my own notions, which I have for several years now taught. A very precise account is given in Cunningham's Text-book of Anatomy. The maxilla is there stated to be developed in the connective tissue around the oral cavity of the embryo from centres which are not preceded by cartilage, of uncertain number, as early fusion takes place between them. -

Morfofunctional Structure of the Skull
N.L. Svintsytska V.H. Hryn Morfofunctional structure of the skull Study guide Poltava 2016 Ministry of Public Health of Ukraine Public Institution «Central Methodological Office for Higher Medical Education of MPH of Ukraine» Higher State Educational Establishment of Ukraine «Ukranian Medical Stomatological Academy» N.L. Svintsytska, V.H. Hryn Morfofunctional structure of the skull Study guide Poltava 2016 2 LBC 28.706 UDC 611.714/716 S 24 «Recommended by the Ministry of Health of Ukraine as textbook for English- speaking students of higher educational institutions of the MPH of Ukraine» (minutes of the meeting of the Commission for the organization of training and methodical literature for the persons enrolled in higher medical (pharmaceutical) educational establishments of postgraduate education MPH of Ukraine, from 02.06.2016 №2). Letter of the MPH of Ukraine of 11.07.2016 № 08.01-30/17321 Composed by: N.L. Svintsytska, Associate Professor at the Department of Human Anatomy of Higher State Educational Establishment of Ukraine «Ukrainian Medical Stomatological Academy», PhD in Medicine, Associate Professor V.H. Hryn, Associate Professor at the Department of Human Anatomy of Higher State Educational Establishment of Ukraine «Ukrainian Medical Stomatological Academy», PhD in Medicine, Associate Professor This textbook is intended for undergraduate, postgraduate students and continuing education of health care professionals in a variety of clinical disciplines (medicine, pediatrics, dentistry) as it includes the basic concepts of human anatomy of the skull in adults and newborns. Rewiewed by: O.M. Slobodian, Head of the Department of Anatomy, Topographic Anatomy and Operative Surgery of Higher State Educational Establishment of Ukraine «Bukovinian State Medical University», Doctor of Medical Sciences, Professor M.V. -

Comparison of Greater Palatine Nerve Block with Intravenous Fentanyl for Postoperative Analgesia Following Palatoplasty in Children
Jemds.com Original Research Article Comparison of Greater Palatine Nerve Block with Intravenous Fentanyl for Postoperative Analgesia Following Palatoplasty in Children Amol Singam1, Saranya Rallabhandi2, Tapan Dhumey3 1Department of Anaesthesiology, JNMC, DMIMS, Sawangi Meghe, Wardha Maharashtra, India. 2Department of Anaesthesiology, JNMC, DMIMS, Sawangi Meghe, Wardha, Maharashtra, India. 3Department of Anaesthesiology, JNMC, DMIMS, Sawangi Meghe, Wardha, Maharashtra, India. ABSTRACT BACKGROUND Good pain relief after palatoplasty is important as inadequate analgesia with vigorous Corresponding Author: cry leads to wound dehiscence, removal of sutures and extra nursing care. Decrease Dr. Saranya Rallabhandi, in oxygen requirement and cardio-respiratory demand occur with good pain relief Assisstant Professor, and also promotes early recovery. Preoperative opioids have concerns like sedation, Department of Anesthesiology, AVBRH, DMIMS (DU), Sawangi Meghe, respiratory depression and airway compromise. Greater palatine nerve block with Wardha- 442001, Maharashtra, India. bupivacaine is safe and effective without the risk of respiratory depression. The study E-mail: [email protected] was done to compare pain relief postoperatively with intravenous fentanyl and greater palatine nerve block in children following palatoplasty. DOI: 10.14260/jemds/2020/549 METHODS How to Cite This Article: 80 children of ASA I & II, between 1 to 7 years were included and allocated into two Singam A, Rallabhandi S, Dhumey T. Comparison of greater palatine nerve block groups of 40 each. Analgesic medication was given preoperatively after induction of with intravenous fentanyl for postoperative general anaesthesia, children in Group B received greater palatine nerve block with analgesia following palatoplasty in -1 2 mL 0.25% inj. Bupivacaine (1 mL on each side) and Group F received 2 μg Kg I.V. -

MAXILLARY FRACTURES Is Mandatory and May Include Both Plain Films and a Computed Tomographic (CT) the Maxilla Forms the Largest Component of Scan
Radiographic evaluation of the fracture MAXILLARY FRACTURES is mandatory and may include both plain films and a computed tomographic (CT) The maxilla forms the largest component of scan. The CT scan has now essentially the middle third of the facial skeleton. The maxil- replaced plain films as the Ògold standardÓ la is a key bone in the midface that is closely asso- in both evaluation and treatment planning. ciated with adjacent bones providing structural If physical findings and plain films are not support between the cranial base and the occlusal suggestive of a zygomatic fracture, the eval- plane. Fractures of the maxilla occur less fre- uation may end here. However, if they do quently than those of the mandible or nose due to suggest fracture, a coronal and axial CT the strong structural support of this bone. The scan should be obtained. The CT scan will midface consists of alternating thick and thin sec- accurately reveal the extent of orbital tions of bone that are capable of resisting signifi- involvement, as well as degree of displace- cant force. This structurally strong bone provides ment of the fractures. This study is vital for protection for the globes and brain, projection of planning the operative approach. the midface, and support for occlusion. Reestablishing continuity of these buttresses is the Historically, closed reduction was the foundation on which maxillary fracture treatment method of choice for nearly all zygomatic is based. fractures. Multiple methods were employed, but most involved simply exert- Renee LeFort (1901) provided the earliest clas- ing pressure underneath the malar emi- sification system of maxillary fractures. -

Results Description of the SKULLS. the Overall Size of Both Skulls Was Considered to Be Within Normal Limits for Their Ethnic
Ossification Defects and Craniofacial Morphology In Incomplete Forms of Mandibulofacial Dysostosis A Description of Two Dry Skulls ERIK DAHL, D.D.S., DR. ODONT. ARNE BJORK, D.D.S., ODONT. DR. Copenhagen, Denmark The morphology of two East Indian dry skulls exhibiting anomalies which were suggested to represent incomplete forms of mandibulofacial dysostosis is described. Obvious although minor ossification anomalies were found localized to the temporal, sphenoid, the zygomatic, the maxillary and the mandibular bones. The observations substantiate the concept of the regional and bilateral nature of this malformation syndrome. Bilateral orbital deviations, hypoplasia of the malar bones, and incomplete zygomatic arches appear to be hard tissue aberrations which may be helpful in exami- nation for subclinical carrier status. Changes in mandibular morphology seem to be less distinguishing features in incomplete or abortive types of mandibulofacial dysostosis. KEY WORDS craniofacial problems, mandible, mandibulofacial dysostosis, maxilla, sphenoid bone, temporal bone, zygomatic bone Mandibulofacial dysostosis (MFD) often roentgencephalometric examinations were results in the development of a characteristic made of the skulls, and tomograms were ob- facial disfigurement with considerable simi- tained of the internal and middle ear. Com- larity between affected individuals. However, parisons were made with normal adult skulls the symptoms may vary highly in respect to and with an adult skull exhibiting the char- type and degree, and both incomplete and acteristics of MFD. All of the skulls were from abortive forms of the syndrome have been the same ethnic group. ' reported in the literature (Franceschetti and Klein, 1949; Moss et al., 1964; Rogers, 1964). Results In previous papers, we have shown the DEsCRIPTION OF THE SKULLS. -

Analysis of Facial Skeletal Morphology: Nasal Bone, Maxilla, and Mandible
Hindawi BioMed Research International Volume 2021, Article ID 5599949, 9 pages https://doi.org/10.1155/2021/5599949 Research Article Analysis of Facial Skeletal Morphology: Nasal Bone, Maxilla, and Mandible Han-Sheng Chen ,1 Szu-Yu Hsiao ,2,3 and Kun-Tsung Lee 4,5 1Dental Department, Kaohsiung Municipal Siao-gang Hospital, Kaohsiung, Taiwan 2School of Dental Medicine, Kaohsiung Medical University, Kaohsiung, Taiwan 3Department of Dentistry for Child and Special Needs, Kaohsiung Medical University Hospital, Kaohsiung, Taiwan 4Division of Clinical Dentistry, Department of Dentistry, Kaohsiung Medical University Hospital, Kaohsiung, Taiwan 5Department of Oral Hygiene, College of Dental Science, Kaohsiung Medical University, Kaohsiung, Taiwan Correspondence should be addressed to Kun-Tsung Lee; [email protected] Received 12 February 2021; Revised 29 March 2021; Accepted 4 May 2021; Published 25 May 2021 Academic Editor: Michael YC Chen Copyright © 2021 Han-Sheng Chen et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. The growth and development of facial bones are closely related to each other. The present study investigated the differences in the nasomaxillary and mandibular morphology among different skeletal patterns. Cephalograms of 240 participants were divided into 3 groups based on the skeletal pattern (Class I, Class II, and Class III). The dimensions of nasomaxilla (nasal bone length, nasal ridge length, nasal depth, palatal length, and maxillary height) and mandible (condylar length, ramus length, body length, symphysis length, and entire mandibular length) were measured. One-way analysis of variance and Pearson’s correlation test were used for statistical analysis. -

MRI-Based Assessment of Masticatory Muscle Changes in TMD Patients After Whiplash Injury
Journal of Clinical Medicine Article MRI-Based Assessment of Masticatory Muscle Changes in TMD Patients after Whiplash Injury Yeon-Hee Lee 1,* , Kyung Mi Lee 2 and Q-Schick Auh 1 1 Department of Orofacial Pain and Oral Medicine, Kyung Hee University Dental Hospital, #613 Hoegi-dong, Dongdaemun-gu, Seoul 02447, Korea; [email protected] 2 Department of Radiology, Kyung Hee University College of Medicine, Kyung Hee University Hospital, #26 Kyunghee-daero, Dongdaemun-gu, Seoul 02447, Korea; [email protected] * Correspondence: [email protected]; Tel.: +82-2-958-9409; Fax: +82-2-968-0588 Abstract: Objective: to investigate the change in volume and signal in the masticatory muscles and temporomandibular joint (TMJ) of patients with temporomandibular disorder (TMD) after whiplash injury, based on magnetic resonance imaging (MRI), and to correlate them with other clinical parameters. Methods: ninety patients (64 women, 26 men; mean age: 39.36 ± 15.40 years), including 45 patients with symptoms of TMD after whiplash injury (wTMD), and 45 age- and sex- matched controls with TMD due to idiopathic causes (iTMD) were included. TMD was diagnosed using the study diagnostic criteria for TMD Axis I, and MRI findings of the TMJ and masticatory muscles were investigated. To evaluate the severity of TMD pain and muscle tenderness, we used a visual analog scale (VAS), palpation index (PI), and neck PI. Results: TMD indexes, including VAS, PI, and neck PI were significantly higher in the wTMD group. In the wTMD group, muscle tenderness was highest in the masseter muscle (71.1%), and muscle tenderness in the temporalis (60.0%), lateral pterygoid muscle (LPM) (22.2%), and medial pterygoid muscle (15.6%) was significantly more frequent than that in the iTMD group (all p < 0.05). -

Tinnitus and Temporomandibular Joint Disorder Subtypes
TINNITUS AND TEMPOROMANDIBULAR JOINT DISORDER SUBTYPES SUSEE PRIYANKA RAVURI A thesis Submitted in partial fulfillment of the requirements for the degree of MASTER OF SCIENCE IN DENTISTRY University of Washington 2017 Committee Edmond L. Truelove Peggy Lee Lloyd A. Mancl Program Authorized to Offer Degree: Oral Medicine 1 © Copyright 2017 Susee Priyanka Ravuri 2 University of Washington ABSTRACT Tinnitus And Temporomandibular Joint Disorder Subtypes Susee Priyanka Ravuri Edmond L. Truelove B.S., D.D.S., M.S.D. Oral Medicine OBJECTIVE: The purpose of this study was to assess the prevalence of tinnitus within a TMD population and to determine an association between the presence of tinnitus and type of TMD diagnoses. METHODS: A secondary data analysis was performed using data from ‘Research Diagnostic Criteria for Temporomandibular Disorders (RDC/TMD) baseline (Validation project) study and follow up (Impact project) study. Self-reported questionnaires for reporting tinnitus and medical history and gold standard diagnoses after clinical examination were used. Log-binomial regression was used to compute risk ratios for tinnitus by TMD subtype and adjusted for patient characteristics. All statistical analysis was performed using SAS 9.3 software (SAS Institute), and a two-sided significance level of 0.05 to determined statistical significance (p<0.05). RESULTS: At baseline, 614 subjects met required criteria for TMD diagnosis. Prevalence of tinnitus within sample was 41% (253 of 614). Approximately 80% of TMD subjects received a MPD diagnosis. Tinnitus frequency in the MPD group was 48% (238/495) while subjects without MPD diagnosis the rate of tinnitus was 13% (15 of 119). Using log-binomial regression analysis, the risk ratio for tinnitus was calculated. -
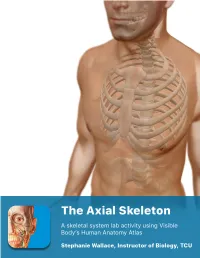
Lab Manual Axial Skeleton Atla
1 PRE-LAB EXERCISES When studying the skeletal system, the bones are often sorted into two broad categories: the axial skeleton and the appendicular skeleton. This lab focuses on the axial skeleton, which consists of the bones that form the axis of the body. The axial skeleton includes bones in the skull, vertebrae, and thoracic cage, as well as the auditory ossicles and hyoid bone. In addition to learning about all the bones of the axial skeleton, it is also important to identify some significant bone markings. Bone markings can have many shapes, including holes, round or sharp projections, and shallow or deep valleys, among others. These markings on the bones serve many purposes, including forming attachments to other bones or muscles and allowing passage of a blood vessel or nerve. It is helpful to understand the meanings of some of the more common bone marking terms. Before we get started, look up the definitions of these common bone marking terms: Canal: Condyle: Facet: Fissure: Foramen: (see Module 10.18 Foramina of Skull) Fossa: Margin: Process: Throughout this exercise, you will notice bold terms. This is meant to focus your attention on these important words. Make sure you pay attention to any bold words and know how to explain their definitions and/or where they are located. Use the following modules to guide your exploration of the axial skeleton. As you explore these bones in Visible Body’s app, also locate the bones and bone markings on any available charts, models, or specimens. You may also find it helpful to palpate bones on yourself or make drawings of the bones with the bone markings labeled. -

Imaging of Mandibular Fractures: a Pictorial Review
Nardi et al. Insights into Imaging (2020) 11:30 https://doi.org/10.1186/s13244-020-0837-0 Insights into Imaging EDUCATIONAL REVIEW Open Access Imaging of mandibular fractures: a pictorial review Cosimo Nardi1, Chiara Vignoli1, Michele Pietragalla1, Paolina Tonelli1, Linda Calistri1, Lorenzo Franchi2,3, Lorenzo Preda4,5* and Stefano Colagrande1 Abstract Mandibular fractures are among the most common maxillofacial fractures observed in emergency rooms and are mainly caused by road accidents. The clinical features of mandibular fractures include malocclusion and loss of mandibular function. Panoramic radiography is usually limited to isolated lesions, whereas computed tomography is the tool of choice for all other facial traumatic events. No reference standard classification system for the different types of mandibular fractures is defined. Therapeutic options include a conservative approach or surgical treatment based on the anatomic area and the severity of fracture. The main purpose of this pictorial review is to illustrate a practical description of the pathophysiology of mandibular fractures and describe both the imaging techniques to recognise them and the therapeutic indications. Keywords: Mandible, Condyle, Fracture, Trauma, Panoramic radiography Key points maxillofacial fractures varies according to geographical Mandibular fractures represent two thirds of all areas and socio-economic factors. The most common maxillofacial fractures. causes of maxillofacial fractures are road traffic accidents X-ray films, including panoramic radiography, are (40–42%), falls, assaults, sports, and work injuries [3]. usually limited to mild traumatic events. The average age of patients with mandibular fracture is Computed tomography is the tool of choice for the 38 years for men and 40 years for women [4]. -

MBB: Head & Neck Anatomy
MBB: Head & Neck Anatomy Skull Osteology • This is a comprehensive guide of all the skull features you must know by the practical exam. • Many of these structures will be presented multiple times during upcoming labs. • This PowerPoint Handout is the resource you will use during lab when you have access to skulls. Mind, Brain & Behavior 2021 Osteology of the Skull Slide Title Slide Number Slide Title Slide Number Ethmoid Slide 3 Paranasal Sinuses Slide 19 Vomer, Nasal Bone, and Inferior Turbinate (Concha) Slide4 Paranasal Sinus Imaging Slide 20 Lacrimal and Palatine Bones Slide 5 Paranasal Sinus Imaging (Sagittal Section) Slide 21 Zygomatic Bone Slide 6 Skull Sutures Slide 22 Frontal Bone Slide 7 Foramen RevieW Slide 23 Mandible Slide 8 Skull Subdivisions Slide 24 Maxilla Slide 9 Sphenoid Bone Slide 10 Skull Subdivisions: Viscerocranium Slide 25 Temporal Bone Slide 11 Skull Subdivisions: Neurocranium Slide 26 Temporal Bone (Continued) Slide 12 Cranial Base: Cranial Fossae Slide 27 Temporal Bone (Middle Ear Cavity and Facial Canal) Slide 13 Skull Development: Intramembranous vs Endochondral Slide 28 Occipital Bone Slide 14 Ossification Structures/Spaces Formed by More Than One Bone Slide 15 Intramembranous Ossification: Fontanelles Slide 29 Structures/Apertures Formed by More Than One Bone Slide 16 Intramembranous Ossification: Craniosynostosis Slide 30 Nasal Septum Slide 17 Endochondral Ossification Slide 31 Infratemporal Fossa & Pterygopalatine Fossa Slide 18 Achondroplasia and Skull Growth Slide 32 Ethmoid • Cribriform plate/foramina