The Development of the Human Maxilla, Vomer, and Paraseptal Cartilages
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Download PDF File
Folia Morphol. Vol. 78, No. 2, pp. 331–343 DOI: 10.5603/FM.a2018.0084 O R I G I N A L A R T I C L E Copyright © 2019 Via Medica ISSN 0015–5659 journals.viamedica.pl Morphometric evaluation and surgical implications of the infraorbital groove, canal and foramen on cone-beam computed tomography and a review of literature İ. Bahşi1, M. Orhan1, P. Kervancioğlu1, E.D. Yalçin2 1Department of Anatomy, Faculty of Medicine, Gaziantep University, Gaziantep, Turkey 2Department of Dentomaxillofacial Radiology, Faculty of Dentistry, Gaziantep University, Gaziantep, Turkey [Received: 25 June 2018; Accepted: 8 August 2018] Background: The purpose of this study is to evaluate the anatomy, morphometry, and variations of infraorbital groove (IOG), infraorbital canal (IOC) and infraorbital foramen (IOF) on the cone-beam computed tomography (CBCT) images and to investigate their relations with surrounding structures. Methods: IOG, IOC and IOF were evaluated retrospectively in CBCT images of 75 female (F) and 75 male (M) cases with a range of 18–65 years (F: 37.62 ± ± 13.55, M: 37.53 ± 15.87) by Planmeca Romexis programme. IOG, IOC and IOF were examined bilaterally (300 sides) in the cases. The 13 parameters were measured on these images in axial, sagittal and coronal planes. Results: There was a very weak positive correlation between the age and the angle between IOC and IOG (p = 0.015, r = 0.198), there was a weak positive correlation between the age and skin thickness (p = 0.001, r = 0.281), and there was no correlation between the age and other parameters. -

MAXILLARY FRACTURES Is Mandatory and May Include Both Plain Films and a Computed Tomographic (CT) the Maxilla Forms the Largest Component of Scan
Radiographic evaluation of the fracture MAXILLARY FRACTURES is mandatory and may include both plain films and a computed tomographic (CT) The maxilla forms the largest component of scan. The CT scan has now essentially the middle third of the facial skeleton. The maxil- replaced plain films as the Ògold standardÓ la is a key bone in the midface that is closely asso- in both evaluation and treatment planning. ciated with adjacent bones providing structural If physical findings and plain films are not support between the cranial base and the occlusal suggestive of a zygomatic fracture, the eval- plane. Fractures of the maxilla occur less fre- uation may end here. However, if they do quently than those of the mandible or nose due to suggest fracture, a coronal and axial CT the strong structural support of this bone. The scan should be obtained. The CT scan will midface consists of alternating thick and thin sec- accurately reveal the extent of orbital tions of bone that are capable of resisting signifi- involvement, as well as degree of displace- cant force. This structurally strong bone provides ment of the fractures. This study is vital for protection for the globes and brain, projection of planning the operative approach. the midface, and support for occlusion. Reestablishing continuity of these buttresses is the Historically, closed reduction was the foundation on which maxillary fracture treatment method of choice for nearly all zygomatic is based. fractures. Multiple methods were employed, but most involved simply exert- Renee LeFort (1901) provided the earliest clas- ing pressure underneath the malar emi- sification system of maxillary fractures. -

Atlas of the Facial Nerve and Related Structures
Rhoton Yoshioka Atlas of the Facial Nerve Unique Atlas Opens Window and Related Structures Into Facial Nerve Anatomy… Atlas of the Facial Nerve and Related Structures and Related Nerve Facial of the Atlas “His meticulous methods of anatomical dissection and microsurgical techniques helped transform the primitive specialty of neurosurgery into the magnificent surgical discipline that it is today.”— Nobutaka Yoshioka American Association of Neurological Surgeons. Albert L. Rhoton, Jr. Nobutaka Yoshioka, MD, PhD and Albert L. Rhoton, Jr., MD have created an anatomical atlas of astounding precision. An unparalleled teaching tool, this atlas opens a unique window into the anatomical intricacies of complex facial nerves and related structures. An internationally renowned author, educator, brain anatomist, and neurosurgeon, Dr. Rhoton is regarded by colleagues as one of the fathers of modern microscopic neurosurgery. Dr. Yoshioka, an esteemed craniofacial reconstructive surgeon in Japan, mastered this precise dissection technique while undertaking a fellowship at Dr. Rhoton’s microanatomy lab, writing in the preface that within such precision images lies potential for surgical innovation. Special Features • Exquisite color photographs, prepared from carefully dissected latex injected cadavers, reveal anatomy layer by layer with remarkable detail and clarity • An added highlight, 3-D versions of these extraordinary images, are available online in the Thieme MediaCenter • Major sections include intracranial region and skull, upper facial and midfacial region, and lower facial and posterolateral neck region Organized by region, each layered dissection elucidates specific nerves and structures with pinpoint accuracy, providing the clinician with in-depth anatomical insights. Precise clinical explanations accompany each photograph. In tandem, the images and text provide an excellent foundation for understanding the nerves and structures impacted by neurosurgical-related pathologies as well as other conditions and injuries. -

Three-Dimensional Radiographic Evaluation of the Malar Bone Engagement Available for Ideal Zygomatic Implant Placement
Article Three-Dimensional Radiographic Evaluation of the Malar Bone Engagement Available for Ideal Zygomatic Implant Placement Gerardo Pellegrino 1,* , Francesco Grande 2 , Agnese Ferri 1, Paolo Pisi 3, Maria Giovanna Gandolfi 4 and Claudio Marchetti 1 1 Oral and Maxillofacial Surgery Unit, Department of Biomedical and Neuromotor Sciences, University of Bologna, 40125 Bologna, Italy; [email protected] (A.F.); [email protected] (C.M.) 2 Oral Surgery Unit, Dental School, Department of Biomedical and Neuromotor Sciences, University of Bologna, 40125 Bologna, Italy; [email protected] 3 Dental Radiology Unit, Dental School, Department of Biomedical and Neuromotor Sciences, University of Bologna, 40125 Bologna, Italy; [email protected] 4 Medical-technical Science, Dental School, Department of Biomedical and Neuromotor Sciences, University of Bologna, 40125 Bologna, Italy; mgiovanna.gandolfi@unibo.it * Correspondence: [email protected]; Tel.: +39-051-208-8157 Received: 11 June 2020; Accepted: 21 July 2020; Published: 22 July 2020 Abstract: Zygomatic implant rehabilitation is a challenging procedure that requires an accurate prosthetic and implant plan. The aim of this study was to evaluate the malar bone available for three-dimensional zygomatic implant placement on the possible trajectories exhibiting optimal occlusal emergence. After a preliminary analysis on 30 computed tomography (CT) scans of dentate patients to identify the ideal implant emergencies, we used 80 CT scans of edentulous patients to create two sagittal planes representing the possible trajectories of the anterior and posterior zygomatic implants. These planes were rotated clockwise on the ideal emergence points and three different hypothetical implant trajectories per zygoma were drawn for each slice. -

A Review of the Mandibular and Maxillary Nerve Supplies and Their Clinical Relevance
AOB-2674; No. of Pages 12 a r c h i v e s o f o r a l b i o l o g y x x x ( 2 0 1 1 ) x x x – x x x Available online at www.sciencedirect.com journal homepage: http://www.elsevier.com/locate/aob Review A review of the mandibular and maxillary nerve supplies and their clinical relevance L.F. Rodella *, B. Buffoli, M. Labanca, R. Rezzani Division of Human Anatomy, Department of Biomedical Sciences and Biotechnologies, University of Brescia, V.le Europa 11, 25123 Brescia, Italy a r t i c l e i n f o a b s t r a c t Article history: Mandibular and maxillary nerve supplies are described in most anatomy textbooks. Accepted 20 September 2011 Nevertheless, several anatomical variations can be found and some of them are clinically relevant. Keywords: Several studies have described the anatomical variations of the branching pattern of the trigeminal nerve in great detail. The aim of this review is to collect data from the literature Mandibular nerve and gives a detailed description of the innervation of the mandible and maxilla. Maxillary nerve We carried out a search of studies published in PubMed up to 2011, including clinical, Anatomical variations anatomical and radiological studies. This paper gives an overview of the main anatomical variations of the maxillary and mandibular nerve supplies, describing the anatomical variations that should be considered by the clinicians to understand pathological situations better and to avoid complications associated with anaesthesia and surgical procedures. # 2011 Elsevier Ltd. -
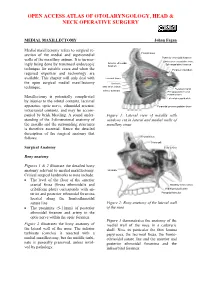
MEDIAL MAXILLECTOMY Johan Fagan
OPEN ACCESS ATLAS OF OTOLARYNGOLOGY, HEAD & NECK OPERATIVE SURGERY MEDIAL MAXILLECTOMY Johan Fagan Medial maxillectomy refers to surgical re- section of the medial and superomedial Frontal sinus walls of the maxillary antrum. It is increas- Posterior ethmoidal foramen Orbital process palatine bone Anterior ethmoidal Sphenopalatine foramen ingly being done by transnasal endoscopic foramen technique for suitable cases and when the Foramen rotundum required expertise and technology are available. This chapter will only deal with Lacrimal fossa the open surgical medial maxillectomy Uncinate Max sinus ostium technique. Pterygoid canal Inferior turbinate Pterygopalatine canal Palatine bone Maxillectomy is potentially complicated Lateral pterygoid plate by injuries to the orbital contents, lacrimal apparatus, optic nerve, ethmoidal arteries, Pyramidal process palatine bone intracranial contents, and may be accom- panied by brisk bleeding. A sound under- Figure 1: Lateral view of maxilla with standing of the 3-dimensional anatomy of windows cut in lateral and medial walls of the maxilla and the surrounding structures maxillary sinus is therefore essential. Hence the detailed description of the surgical anatomy that follows. Frontal sinus Crista galli Surgical Anatomy Sella turcica Bony anatomy Figures 1 & 2 illustrate the detailed bony anatomy relevant to medial maxillectomy. Uncinate Critical surgical landmarks to note include: • The level of the floor of the anterior cranial fossa (fovea ethmoidalis and Maxillary sinus ostium cribriform plate) corresponds with an- Medial pterygoid plate terior and posterior ethmoidal foramina Pterygoid hamulus located along the frontoethmoidal suture line Figure 2: Bony anatomy of the lateral wall • The proximity (5-11mm) of posterior of the nose ethmoidal foramen and artery to the optic nerve within the optic foramen Figure 3 demonstrates the anatomy of the Figure 2 illustrates the bony anatomy of medial wall of the nose in a cadaveric the lateral wall of the nose. -

A STUDY on POSITION of INFRAORBITAL FORAMEN Shaik Hussain Saheb 1, Shruthi B.N *2, Pavan P Havaldar 3
International Journal of Anatomy and Research, Int J Anat Res 2017, Vol 5(3.2):4257-60. ISSN 2321-4287 Original Research Article DOI: https://dx.doi.org/10.16965/ijar.2017.300 A STUDY ON POSITION OF INFRAORBITAL FORAMEN Shaik Hussain Saheb 1, Shruthi B.N *2, Pavan P Havaldar 3. 1 Department of Anatomy, JJM Medical College, Davangere, Karnataka, India. *2 Department of Anatomy, Rajarajeswari Medical College and hospital, Bangalore, Karnataka, India. 3 Department of Anatomy, Gadag institute of medical sciences, Gadag, Karnataka, India. ABSTRACT Background: The infraorbital foramen is located on the maxillary bone about 1 cm inferior to the infraorbital margin. The infraorbital nerve and vessels are transmitted through this foramen. The infraorbital nerve, the continuation of the maxillary or second division of the trigeminal nerve, is solely a sensory nerve. It traverses the inferior orbital fissure into the inferior orbital canal and emerges onto the face at the infraorbital foramen. It divides into several branches that innervate the skin and the mucous membrane of the midface, such as the lower eyelid, cheek, lateral aspect of the nose, upper lip, and the labial gum. Materials and Methods: Total 300 skulls were used for this study, the following mesearements were recorded, mean distance between the infra orbital foramen and the infra orbital margin on right and left side and average of it. The mean distance between the infra orbital foramen and the piriform aperature on right and left side measured and average of it also recorded. The mean distance between infra orbital foramen and the anterior nasal spine on right and left side measured. -

Evaluation of the Infraorbital Complex by Direct Morphometry
Romanian Journal of Oral Rehabilitation Vol. 12, No. 3, July - September 2020 EVALUATION OF THE INFRAORBITAL COMPLEX BY DIRECT MORPHOMETRY Maria Justina Roxana Vîrlan 1, Cătălina Murariu-Măgureanu 2, Simona Andreea Moraru 1, Vlad Iliescu 3, Ovidiu Romulus Gherghiţă 4, Vanda Roxana Nimigean 1, Ovidiu Muşat 5, Victor Nimigean 6 1 Oral Rehabilitation Department, Faculty of Dental Medicine, Carol Davila University of Medicine and Pharmacy, Bucharest, Romania; 2 Complete Denture Prosthodontics, Faculty of Dental Medicine, Carol Davila University of Medicine and Pharmacy, Bucharest, Romania; 3 Private Dental Practice, Bucharest, Romania; 4 PhD student, Faculty of Dental Medicine, Carol Davila University of Medicine and Pharmacy, Bucharest, Romania; 5 Clinical Department of Ophthalmology, Dr. Carol Davila Central Military Emergency University Hospital, Bucharest, Romania; 6 Anatomy Department, Faculty of Dental Medicine, Carol Davila University of Medicine and Pharmacy, Bucharest, Romania. Corresponding author: Vanda Roxana Nimigean, Associate Professor, Head of Oral Rehabilitation Department, Faculty of Dental Medicine, Carol Davila University of Medicine and Pharmacy, Bucharest, Romania, No 17-23 Calea Plevnei Street. E-mail: [email protected] ABSTRACT The infraorbital complex belongs to the maxillary bone and it is an important landmark in dentistry, especially for loco-regional anaesthesia. For this study, bilateral measurements of the components of the infraorbital complex on twenty dry dentated human skulls were performed. Results: the average length of the the infraorbital groove (IOG) was 12.55 mm and the average length of the infraorbital canal (IOC) was 12.92 mm. Regarding the infraorbital foramen (IOF), its shape was oval in 65% of the cases and the average distance between the infraorbital foramen and the inferior orbital margin (IOF-IOM distance) was 7.87 mm. -

INFERIOR MAXILLECTOMY Johan Fagan
OPEN ACCESS ATLAS OF OTOLARYNGOLOGY, HEAD & NECK OPERATIVE SURGERY INFERIOR MAXILLECTOMY Johan Fagan Tumours of the hard palate and superior Figure 2 illustrates the bony anatomy of alveolus may be resected by inferior the lateral wall of the nose. The inferior maxillectomy (Figure 1). A Le Fort 1 turbinate (concha) may be resected with osteotomy may also be used as an inferior maxillectomy, but the middle tur- approach to e.g. angiofibromas and the binate is preserved. nasopharynx. Frontal sinus Posterior ethmoidal foramen Orbital process palatine bone Anterior ethmoidal Sphenopalatine foramen foramen Foramen rotundum Lacrimal fossa Uncinate Max sinus ostium Pterygoid canal Inferior turbinate Pterygopalatine canal Palatine bone Lateral pterygoid plate Figure 1: Bilateral inferior maxillectomy Pyramidal process palatine bone A sound understanding of the 3-dimen- Figure 2: Lateral view of maxilla with sional anatomy of the maxilla and the windows cut in lateral and medial walls of surrounding structures is essential to do the maxillary sinus operation safely. Hence the detailed description of the relevant surgical anatomy that follows. Frontal sinus Crista galli Surgical Anatomy Sella turcica Bony anatomy Figures 2, 3 & 4 illustrate the detailed bony anatomy relevant to maxillectomy. Uncinate Critical surgical landmarks to note include: • The floor of the anterior cranial fossa (fovea ethmoidalis and cribriform Maxillary sinus ostium plate) corresponds with anterior and Medial pterygoid plate posterior ethmoidal foramina located, Pterygoid -

Evaluation of Maxillary Bone Dimensions in Specific Areas for Removable Dentures
https://doi.org/10.5272/jimab.2017232.1527 Journal of IMAB Journal of IMAB - Annual Proceeding (Scientific Papers). 2017 Apr-Jun;23(2): ISSN: 1312-773X https://www.journal-imab-bg.org Original article EVALUATION OF MAXILLARY BONE DIMENSIONS IN SPECIFIC AREAS FOR REMOVABLE DENTURES Original Articles Dobromira Shopova1, Tanya Bozhkova1, Dian Slavchev1, Spas Muletarov2, Zdravka Ivanova3, Elena Bozhikova2 1) Department of Prosthetic Dentistry, Faculty of Dental Medicine, Medical University - Plovdiv, Bulgaria; 2) Department of Anatomy, Histology and Embryology, Medical University - Plovdiv, Bulgaria; 3) Department of Plant Physiology and Molecular Biology, Plovdiv University - Plovdiv, Bulgaria. ABSTRACT similar studies for quantitative evaluation module of elas- Background: Removable prosthetics is a big part of ticity and hardness in different anatomical regions, and Prosthetic Dentistry. Prosthetic field is very important for they were established middle level in frontal area and low successful treatment with partial or complete dentures. in distal zone [3, 4, 5]. The cortical density can be meas- Maxillary bone is covered with soft tissues, but its anatomy ured by Hounsfield units (HU). The investigations proved is essential for retention, chewing stability and comfort of various levels in cortical and cancellous bone densities. the patients. Maxillary tuberosity showed the lowest level [6]. Com- Purpose: The study’s aim was to evaluate the dimen- puted tomography’s study of the edentulous posterior max- sions of maxillary bone in specific zones for removable illae showed porous cortical crest or no cortical bone, al- dentures. though the bone densities varied markedly among indi- Methods: Sixteen craniums were measured in 10 dif- viduals [7]. Micro-computed tomography (microCT) is a ferent zones. -

Unilateral Upper and Lower Subtotal Maxillectomy Approaches to The
NEUROSURGERY 46:6 | JUNE 2000 | 1416-1453 DOI: 10.1097/00006123-200006000-00025 Anatomic Report Unilateral Upper and Lower Subtotal Maxillectomy Approaches to the Cranial Base: Downloaded from https://academic.oup.com/neurosurgery/article-abstract/46/6/1416/2925972 by Universidad de Zaragoza user on 02 January 2020 Microsurgical Anatomy Tsutomu Hitotsumatsu, M.D., Ph.D.1, Albert L. Rhoton, Jr., M.D.1 1Department of Neurological Surgery, University of Florida, Gainesville, Florida ABSTRACT OBJECTIVE The relationship of the maxilla, with its thin walls, to the nasal and oral cavities, the orbit, and the infratemporal and pterygopalatine fossae makes it a suitable route for accessing lesions involving both the central and lateral cranial base. In this study, we compared the surgical anatomy and exposure obtained by two unilateral transmaxillary approaches, one directed through an upper subtotal maxillectomy, and the other through a lower subtotal maxillectomy. METHODS Cadaveric specimens examined, with 3 to 40× magnification, provided the material for this study. RESULTS Both upper and lower maxillectomy approaches open a surgical field extending from the ipsilateral internal carotid artery to the contralateral Eustachian tube; however, they differ in the direction of the access and the areas exposed. The lower maxillectomy opens a combination of the transmaxillary, transnasal, and transoral routes to extra- and intradural lesions of the central cranial base. Performing additional osteotomies of the mandibular coronoid process and the sphenoid pterygoid process provides anterolateral access to the lateral cranial base, including the pterygopalatine and infratemporal fossae, and the parapharyngeal space. The upper maxillectomy opens the transmaxillary and transnasal routes to the central cranial base but not the transoral route. -

The Vomer Bone Analysis in Relation to Class Iii Malocclusion Using Three Dimenssional Images Analysis
International Journal of Dental and Health Sciences Original Article Volume 04,Issue 05 THE VOMER BONE ANALYSIS IN RELATION TO CLASS III MALOCCLUSION USING THREE DIMENSSIONAL IMAGES ANALYSIS Ammar Mohi 1, Kadir Beycan2, Şebnem Erçalik Yalçinkaya3 1Postgraduate PhD researcher, Department of Oral and Maxillofacial Radiology, Faculty of Dentistry, Marmara University, Istanbul, Turkey. 2Assistant professor , Department of Orthodontics, Faculty of Dentistry, Marmara University, Istanbul, Turkey. 3Professor, Chairman and Head of Department of Oral and Maxillofacial Radiology, Faculty of Dentistry, Marmara University, Istanbul, Turkey. ABSTRACT: Objective: To evaluate the vomer bone dimenssional outline changes in relation to the midface hypoplasia of a Class III malocclusion by comparing with normal controls using a three dimenssional CBCT images analysis of Mimics 19.0 software. Material and Method: In total of 96 patients images were both Class III malocclusion as study cases and normal occlusion as controls with age between 15 to 30 years old. All patients were classified into three group based on ANB angular value of Steiner’s analysis. The study group were : normal, mild and sever malocclusion type groups. Linear and angular planes were determined by using 13 skeletal points and analysed by using Mimics 19.0 software. All study groups parameters statistically analysed for significant differences and correlation. Results: A high significant differences between the vomer bone anterior variables (P<0.01) followed by vomer posterior variables (P<0.05) in relation to cranial and midfacial measurements with positive correlation. The pattern of vomer bone was shown highly anterior impaction and backward inclination in sever type malocclusion group and male higher than female.