Nbome) Derivatives
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
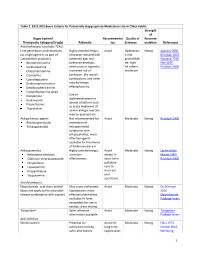
Table 2. 2012 AGS Beers Criteria for Potentially
Table 2. 2012 AGS Beers Criteria for Potentially Inappropriate Medication Use in Older Adults Strength of Organ System/ Recommendat Quality of Recomm Therapeutic Category/Drug(s) Rationale ion Evidence endation References Anticholinergics (excludes TCAs) First-generation antihistamines Highly anticholinergic; Avoid Hydroxyzin Strong Agostini 2001 (as single agent or as part of clearance reduced with e and Boustani 2007 combination products) advanced age, and promethazi Guaiana 2010 Brompheniramine tolerance develops ne: high; Han 2001 Carbinoxamine when used as hypnotic; All others: Rudolph 2008 Chlorpheniramine increased risk of moderate Clemastine confusion, dry mouth, Cyproheptadine constipation, and other Dexbrompheniramine anticholinergic Dexchlorpheniramine effects/toxicity. Diphenhydramine (oral) Doxylamine Use of diphenhydramine in Hydroxyzine special situations such Promethazine as acute treatment of Triprolidine severe allergic reaction may be appropriate. Antiparkinson agents Not recommended for Avoid Moderate Strong Rudolph 2008 Benztropine (oral) prevention of Trihexyphenidyl extrapyramidal symptoms with antipsychotics; more effective agents available for treatment of Parkinson disease. Antispasmodics Highly anticholinergic, Avoid Moderate Strong Lechevallier- Belladonna alkaloids uncertain except in Michel 2005 Clidinium-chlordiazepoxide effectiveness. short-term Rudolph 2008 Dicyclomine palliative Hyoscyamine care to Propantheline decrease Scopolamine oral secretions. Antithrombotics Dipyridamole, oral short-acting* May -

Appendix A: Potentially Inappropriate Prescriptions (Pips) for Older People (Modified from ‘STOPP/START 2’ O’Mahony Et Al 2014)
Appendix A: Potentially Inappropriate Prescriptions (PIPs) for older people (modified from ‘STOPP/START 2’ O’Mahony et al 2014) Consider holding (or deprescribing - consult with patient): 1. Any drug prescribed without an evidence-based clinical indication 2. Any drug prescribed beyond the recommended duration, where well-defined 3. Any duplicate drug class (optimise monotherapy) Avoid hazardous combinations e.g.: 1. The Triple Whammy: NSAID + ACE/ARB + diuretic in all ≥ 65 year olds (NHS Scotland 2015) 2. Sick Day Rules drugs: Metformin or ACEi/ARB or a diuretic or NSAID in ≥ 65 year olds presenting with dehydration and/or acute kidney injury (AKI) (NHS Scotland 2015) 3. Anticholinergic Burden (ACB): Any additional medicine with anticholinergic properties when already on an Anticholinergic/antimuscarinic (listed overleaf) in > 65 year olds (risk of falls, increased anticholinergic toxicity: confusion, agitation, acute glaucoma, urinary retention, constipation). The following are known to contribute to the ACB: Amantadine Antidepressants, tricyclic: Amitriptyline, Clomipramine, Dosulepin, Doxepin, Imipramine, Nortriptyline, Trimipramine and SSRIs: Fluoxetine, Paroxetine Antihistamines, first generation (sedating): Clemastine, Chlorphenamine, Cyproheptadine, Diphenhydramine/-hydrinate, Hydroxyzine, Promethazine; also Cetirizine, Loratidine Antipsychotics: especially Clozapine, Fluphenazine, Haloperidol, Olanzepine, and phenothiazines e.g. Prochlorperazine, Trifluoperazine Baclofen Carbamazepine Disopyramide Loperamide Oxcarbazepine Pethidine -

Modulation by Trace Amine-Associated Receptor 1 of Experimental Parkinsonism, L-DOPA Responsivity, and Glutamatergic Neurotransmission
The Journal of Neuroscience, October 14, 2015 • 35(41):14057–14069 • 14057 Neurobiology of Disease Modulation by Trace Amine-Associated Receptor 1 of Experimental Parkinsonism, L-DOPA Responsivity, and Glutamatergic Neurotransmission Alexandra Alvarsson,1* Xiaoqun Zhang,1* Tiberiu L Stan,1 Nicoletta Schintu,1 Banafsheh Kadkhodaei,2 Mark J. Millan,3 Thomas Perlmann,2,4 and Per Svenningsson1 1Department of Clinical Neuroscience, Center for Molecular Medicine, Karolinska Institutet, SE-17176 Stockholm, Sweden, 2Ludwig Institute for Cancer Research, SE-17177 Stockholm, Sweden, 3Pole of Innovation in Neuropsychiatry, Institut de Recherches Servier, Centre de Recherches de Croissy, Paris 87290, France, and 4Department of Cell and Molecular Biology, Karolinska Institutet, SE-17177 Stockholm, Sweden Parkinson’s disease (PD) is a movement disorder characterized by a progressive loss of nigrostriatal dopaminergic neurons. Restoration of dopamine transmission by L-DOPA relieves symptoms of PD but causes dyskinesia. Trace Amine-Associated Receptor 1 (TAAR1) modulates dopaminergic transmission, but its role in experimental Parkinsonism and L-DOPA responses has been neglected. Here, we report that TAAR1 knock-out (KO) mice show a reduced loss of dopaminergic markers in response to intrastriatal 6-OHDA administra- tion compared with wild-type (WT) littermates. In contrast, the TAAR1 agonist RO5166017 aggravated degeneration induced by intra- striatal6-OHDAinWTmice.Subchronic L-DOPAtreatmentofTAAR1KOmiceunilaterallylesionedwith6-OHDAinthemedialforebrain bundle resulted in more pronounced rotational behavior and dyskinesia than in their WT counterparts. The enhanced behavioral sensitization to L-DOPA in TAAR1 KO mice was paralleled by increased phosphorylation of striatal GluA1 subunits of AMPA receptors. Conversely, RO5166017 counteracted both L-DOPA-induced rotation and dyskinesia as well as AMPA receptor phosphorylation. -

Commonly Prescribed Psychotropic Medications
COMMONLY PRESCRIBED PSYCHOTROPIC MEDICATIONS NAME Generic (Trade) DOSAGE KEY CLINICAL INFORMATION Antidepressant Medications* Start: IR-100 mg bid X 4d then ↑ to 100 mg tid; SR-150 mg qam X 4d then ↑ to 150 mg Contraindicated in seizure disorder because it decreases seizure threshold; stimulating; not good for treating anxiety disorders; second Bupropion (Wellbutrin) bid; XL-150 mg qam X 4d, then ↑ to 300 mg qam. Range: 300-450 mg/d. line TX for ADHD; abuse potential. ¢ (IR/SR), $ (XL) Citalopram (Celexa) Start: 10-20 mg qday,↑10-20 mg q4-7d to 30-40 mg qday. Range: 20-60 mg/d. Best tolerated of SSRIs; very few and limited CYP 450 interactions; good choice for anxious pt. ¢ Duloxetine (Cymbalta) Start: 30 mg qday X 1 wk, then ↑ to 60 mg qday. Range: 60-120 mg/d. More GI side effects than SSRIs; tx neuropathic pain; need to monitor BP; 2nd line tx for ADHD. $ Escitalopram (Lexapro) Start: 5 mg qday X 4-7d then ↑ to 10 mg qday. Range 10-30 mg/d (3X potent vs. Celexa). Best tolerated of SSRIs, very few and limited CYP 450 interactions. Good choice for anxious pt. $ Fluoxetine (Prozac) Start: 10 mg qam X 4-7d then ↑ to 20 mg qday. Range: 20-60 mg/d. More activating than other SSRIs; long half-life reduces withdrawal (t ½ = 4-6 d). ¢ Mirtazapine (Remeron) Start: 15 mg qhs. X 4-7d then ↑ to 30 mg qhs. Range: 30-60 mg/qhs. Sedating and appetite promoting; Neutropenia risk (1 in 1000) so avoid in immunosupressed patients. -
![Selective Labeling of Serotonin Receptors Byd-[3H]Lysergic Acid](https://docslib.b-cdn.net/cover/9764/selective-labeling-of-serotonin-receptors-byd-3h-lysergic-acid-319764.webp)
Selective Labeling of Serotonin Receptors Byd-[3H]Lysergic Acid
Proc. Nati. Acad. Sci. USA Vol. 75, No. 12, pp. 5783-5787, December 1978 Biochemistry Selective labeling of serotonin receptors by d-[3H]lysergic acid diethylamide in calf caudate (ergots/hallucinogens/tryptamines/norepinephrine/dopamine) PATRICIA M. WHITAKER AND PHILIP SEEMAN* Department of Pharmacology, University of Toronto, Toronto, Canada M5S 1A8 Communicated by Philip Siekevltz, August 18,1978 ABSTRACT Since it was known that d-lysergic acid di- The objective in this present study was to improve the se- ethylamide (LSD) affected catecholaminergic as well as sero- lectivity of [3H]LSD for serotonin receptors, concomitantly toninergic neurons, the objective in this study was to enhance using other drugs to block a-adrenergic and dopamine receptors the selectivity of [3HJISD binding to serotonin receptors in vitro by using crude homogenates of calf caudate. In the presence of (cf. refs. 36-38). We then compared the potencies of various a combination of 50 nM each of phentolamine (adde to pre- drugs on this selective [3H]LSD binding and compared these clude the binding of [3HJLSD to a-adrenoceptors), apmo ie, data to those for the high-affinity binding of [3H]serotonin and spiperone (added to preclude the binding of [3H[LSD to (39). dopamine receptors), it was found by Scatchard analysis that the total number of 3H sites went down to 300 fmol/mg, compared to 1100 fmol/mg in the absence of the catechol- METHODS amine-blocking drugs. The IC50 values (concentrations to inhibit Preparation of Membranes. Calf brains were obtained fresh binding by 50%) for various drugs were tested on the binding of [3HLSD in the presence of 50 nM each of apomorphine (A), from the Canada Packers Hunisett plant (Toronto). -

Pharmacology and Toxicology of Amphetamine and Related Designer Drugs
Pharmacology and Toxicology of Amphetamine and Related Designer Drugs U.S. DEPARTMENT OF HEALTH AND HUMAN SERVICES • Public Health Service • Alcohol Drug Abuse and Mental Health Administration Pharmacology and Toxicology of Amphetamine and Related Designer Drugs Editors: Khursheed Asghar, Ph.D. Division of Preclinical Research National Institute on Drug Abuse Errol De Souza, Ph.D. Addiction Research Center National Institute on Drug Abuse NIDA Research Monograph 94 1989 U.S. DEPARTMENT OF HEALTH AND HUMAN SERVICES Public Health Service Alcohol, Drug Abuse, and Mental Health Administration National Institute on Drug Abuse 5600 Fishers Lane Rockville, MD 20857 For sale by the Superintendent of Documents, U.S. Government Printing Office Washington, DC 20402 Pharmacology and Toxicology of Amphetamine and Related Designer Drugs ACKNOWLEDGMENT This monograph is based upon papers and discussion from a technical review on pharmacology and toxicology of amphetamine and related designer drugs that took place on August 2 through 4, 1988, in Bethesda, MD. The review meeting was sponsored by the Biomedical Branch, Division of Preclinical Research, and the Addiction Research Center, National Institute on Drug Abuse. COPYRIGHT STATUS The National Institute on Drug Abuse has obtained permission from the copyright holders to reproduce certain previously published material as noted in the text. Further reproduction of this copyrighted material is permitted only as part of a reprinting of the entire publication or chapter. For any other use, the copyright holder’s permission is required. All other matieral in this volume except quoted passages from copyrighted sources is in the public domain and may be used or reproduced without permission from the Institute or the authors. -

From Sacred Plants to Psychotherapy
From Sacred Plants to Psychotherapy: The History and Re-Emergence of Psychedelics in Medicine By Dr. Ben Sessa ‘The rejection of any source of evidence is always treason to that ultimate rationalism which urges forward science and philosophy alike’ - Alfred North Whitehead Introduction: What exactly is it that fascinates people about the psychedelic drugs? And how can we best define them? 1. Most psychiatrists will define psychedelics as those drugs that cause an acute confusional state. They bring about profound alterations in consciousness and may induce perceptual distortions as part of an organic psychosis. 2. Another definition for these substances may come from the cross-cultural dimension. In this context psychedelic drugs may be recognised as ceremonial religious tools, used by some non-Western cultures in order to communicate with the spiritual world. 3. For many lay people the psychedelic drugs are little more than illegal and dangerous drugs of abuse – addictive compounds, not to be distinguished from cocaine and heroin, which are only understood to be destructive - the cause of an individual, if not society’s, destruction. 4. But two final definitions for psychedelic drugs – and those that I would like the reader to have considered by the end of this article – is that the class of drugs defined as psychedelic, can be: a) Useful and safe medical treatments. Tools that as adjuncts to psychotherapy can be used to alleviate the symptoms and course of many mental illnesses, and 1 b) Vital research tools with which to better our understanding of the brain and the nature of consciousness. Classifying psychedelic drugs: 1,2 The drugs that are often described as the ‘classical’ psychedelics include LSD-25 (Lysergic Diethylamide), Mescaline (3,4,5- trimethoxyphenylathylamine), Psilocybin (4-hydroxy-N,N-dimethyltryptamine) and DMT (dimethyltryptamine). -

Prazosin: Preliminary Report and Comparative Studies with Other
298 BRITISH MEDICAL JOURNAL 1 1 MAY 1974 Prazosin: Preliminary Report and Comparative Studies with Other Antihypertensive Agents Br Med J: first published as 10.1136/bmj.2.5914.298 on 11 May 1974. Downloaded from GORDON S. STOKES, MICHAEL A. WEBER British Medical Journal, 1974, 2, 298-300 bilirubin. One patient had minimal grade 3 hypertensive ocu- lar fundus changes, 10 had grade 1 or 2 changes, and four had normal fundi. Serum crea-tinine concentration was normal in Summary every patient. A diagnosis of uncomplicated essential hypertension was In a group of 14 hypertensive patients a 10-week course made in 12 patients. In two other patients pyelographic of treatment with prazosin 3-7 5 mg/day produced a signifi- evidence of analgesic nephropathy was found. The remaining cant reduction in mean blood pressure without serious side patient had essential hypertension and stable angina pectoris. effects. The fall in diastolic pressure exceeded the response After at least two baseline blood pressure readings, taken mm more to a placebo by 10 Hg or in 9 patients. The average when patients were recumbent and standing, treatment was decrease in diastolic pressure was similar to that produced started with prazosin, 1-mg capsules by mouth, three times or by methyldopa 750 mg/day propranolol 120-160 mg/day daily. Thereafter the patient visited the clinic at intervals of was but the fall in systolic pressure comparatively smaller, one to three weeks. At each visit blood pressure was meas- that the consistent with reported experimental work showing ured to the nearest 5 mm Hg with an Accoson sphygmomo- drug causes vasodilatation. -

Near the Himalayas, from Kashmir to Sikkim, at Altitudes the Catholic Inquisition, and the Traditional Use of These of up to 2700 Meters
Year of edition: 2018 Authors of the text: Marc Aixalà & José Carlos Bouso Edition: Alex Verdaguer | Genís Oña | Kiko Castellanos Illustrations: Alba Teixidor EU Project: New Approaches in Harm Reduction Policies and Practices (NAHRPP) Special thanks to collaborators Alejandro Ponce (in Peyote report) and Eduardo Carchedi (in Kambó report). TECHNICAL REPORT ON PSYCHOACTIVE ETHNOBOTANICALS Volumes I - II - III ICEERS International Center for Ethnobotanical Education Research and Service INDEX SALVIA DIVINORUM 7 AMANITA MUSCARIA 13 DATURA STRAMONIUM 19 KRATOM 23 PEYOTE 29 BUFO ALVARIUS 37 PSILOCYBIN MUSHROOMS 43 IPOMOEA VIOLACEA 51 AYAHUASCA 57 IBOGA 67 KAMBÓ 73 SAN PEDRO 79 6 SALVIA DIVINORUM SALVIA DIVINORUM The effects of the Hierba Pastora have been used by Mazatec Indians since ancient times to treat diseases and for divinatory purposes. The psychoactive compound Salvia divinorum contains, Salvinorin A, is the most potent naturally occurring psychoactive substance known. BASIC INFO Ska Pastora has been used in divination and healing Salvia divinorum is a perennial plant native to the Maza- rituals, similar to psilocybin mushrooms. Maria Sabina tec areas of the Sierra Madre Oriental Mountains of Mexi- told Wasson and Hofmann (the discoverers of its Mazatec co. Its habitat is tropical forests, where it grows between usage) that Salvia divinorum was used in times when the- 300 and 800 meters above sea level. It belongs to the re was a shortage of mushrooms. Some sources that have Lamiaceae family, and is mainly reproduced by cuttings done later feldwork point out that the use of S. divinorum since it rarely produces seeds. may be more widespread than originally believed, even in times when mushrooms were abundant. -

Amphetamine Derivatives Interact with Both Plasma Membrane and Secretory Vesicle Biogenic Amine Transporters
'4 0026-895X/93/061227-05503.00/0 Copyright © by The American Society for Pharmacology and Experimental Therapeutics AIl rights of reproduction in any form reserved. MOLECULAR PHARMACOLOGY, 44:1227-1231 Amphetamine Derivatives Interact with Both Plasma Membrane and Secretory Vesicle Biogenic Amine Transporters SHIMON SCHULDINER, SONIA STEINER-MORDOCH, RODRIGO YELIN, STEPHEN C. WALL, and GARY RUDNICK Departmentof Pharmacology,YaleUniversitySchool of Medicine,New Haven,CT 06510 (S.C.W.,G.R.) andAlexanderSilbermanInstituteof Life Sciences,TheHebrew University,GivatRam, Jerusalem,Israel (S.S.,S.S.-M., R.Y.) ReceivedJune 7, 1993; Accepted September 16, 1993 SUMMARY The interaction of fenfluramine, 3,4-methylenedioxymethamphe- porter, fenfiuramine inhibited serotonin transport and dissipated tamine (MDMA), and p-chloroamphetamine (PCA) with the plate- the transmembrane pH difference (ApH) that drives amine up- let plasma membrane serotonin transporter and the vesicular take. The use of [3H]reserpine-binding measurements to deter- amine transporter were studied using both transport and binding mine drug interaction with the vesicular amine transporter al- measurements. Fenfluramine is apparently a substrate for the lowed assessment of the relative ability of MDMA, PCA, and plasma membrane transporter, and consequently inhibits both fenfluramine to bind to the substrate site of the vesicular trans- serotonin transport and imipramine binding. Moreover, fenflura- porter. These measurements permit a distinction between inhi- mine exchanges with internal [3H]serotonin in a plasma mem- bition of vesicular serotonin transport by directly blocking vesic- brane transporter-mediated reaction that requires NaCI and is ular amine transport and by dissipating ApH. The results indicate blocked by imipramine. These properties are similar to those of that MDMA and fenfluramine inhibit by both mechanisms but MDMA and PCA as previously described. -

The American Journal Of
The American Journal of Psychiatry Residents’ Journal July 2015 Volume 10 Issue 7 Inside IN THIS ISSUE 2 New Formats and New Opportunities: The Time to Get Involved is “Now”! Rajiv Radhakrishnan, M.B.B.S., M.D. 3 Prevention of Posttraumatic Stress Disorder: Predicting Response to Trauma Jennifer H. Harris, M.D. 7 Weight Gain in Patients With Schizophrenia: A Recipe For Timely Intervention Ammar El Sara, M.B.Ch.B. 10 Hyperprolactinemia and Antipsychotics: Update for the Training Psychiatrist Stephanie Pope, M.D. 13 A Clinical Case Conference on Spiritual Growth and Healing Elizabeth S. Stevens, D.O. This issue of the Residents’ Journal features a variety of topics. Jennifer H. Har- ris, M.D., discusses prevention of posttraumatic stress disorder, with an overview 15 Priapism: A Rare but Serious of various responses to trauma. Ammar El Sara, M.B.Ch.B., presents a review of Side Effect of Trazodone clinically applicable evidence-based interventions targeting obesity in schizophre- Kamalika Roy, M.D. nia patients. Stephanie Pope, M.D., examines antipsychotic-induced hyperprolac- 17 Classifying Psychopathology: tinemia, including variables affecting prolactin and clinical implications. Elizabeth Mental Kinds and Natural Kinds S. Stevens, D.O., discusses several psychological, social, and spiritual developmen- Reviewed by Aaron J. Hauptman, tal frameworks in a clinical case conference. Kamalika Roy, M.D., presents a case M.D. of priapism as a side effect of trazodone in a middle-aged patient. Lastly, Aaron J. Hauptman, M.D., offers his review of the book Classifying Psychopathology: Mental 18 Residents’ Resources Kinds and Natural Kinds. Editor-in-Chief Associate Editors Editors Emeriti Rajiv Radhakrishnan, M.B.B.S., M.D. -

Psychedelics in Psychiatry: Neuroplastic, Immunomodulatory, and Neurotransmitter Mechanismss
Supplemental Material can be found at: /content/suppl/2020/12/18/73.1.202.DC1.html 1521-0081/73/1/202–277$35.00 https://doi.org/10.1124/pharmrev.120.000056 PHARMACOLOGICAL REVIEWS Pharmacol Rev 73:202–277, January 2021 Copyright © 2020 by The Author(s) This is an open access article distributed under the CC BY-NC Attribution 4.0 International license. ASSOCIATE EDITOR: MICHAEL NADER Psychedelics in Psychiatry: Neuroplastic, Immunomodulatory, and Neurotransmitter Mechanismss Antonio Inserra, Danilo De Gregorio, and Gabriella Gobbi Neurobiological Psychiatry Unit, Department of Psychiatry, McGill University, Montreal, Quebec, Canada Abstract ...................................................................................205 Significance Statement. ..................................................................205 I. Introduction . ..............................................................................205 A. Review Outline ........................................................................205 B. Psychiatric Disorders and the Need for Novel Pharmacotherapies .......................206 C. Psychedelic Compounds as Novel Therapeutics in Psychiatry: Overview and Comparison with Current Available Treatments . .....................................206 D. Classical or Serotonergic Psychedelics versus Nonclassical Psychedelics: Definition ......208 Downloaded from E. Dissociative Anesthetics................................................................209 F. Empathogens-Entactogens . ............................................................209