Cefoxitin, N-Formimidoyl Thienamycin, Clavulanic
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Cusumano-Et-Al-2017.Pdf
International Journal of Infectious Diseases 63 (2017) 1–6 Contents lists available at ScienceDirect International Journal of Infectious Diseases journal homepage: www.elsevier.com/locate/ijid Rapidly growing Mycobacterium infections after cosmetic surgery in medical tourists: the Bronx experience and a review of the literature a a b c b Lucas R. Cusumano , Vivy Tran , Aileen Tlamsa , Philip Chung , Robert Grossberg , b b, Gregory Weston , Uzma N. Sarwar * a Albert Einstein College of Medicine, Montefiore Medical Center, Bronx, New York, USA b Division of Infectious Diseases, Department of Medicine, Albert Einstein College of Medicine, Montefiore Medical Center, Bronx, New York, USA c Department of Pharmacy, Nebraska Medicine, Omaha, Nebraska, USA A R T I C L E I N F O A B S T R A C T Article history: Background: Medical tourism is increasingly popular for elective cosmetic surgical procedures. However, Received 10 May 2017 medical tourism has been accompanied by reports of post-surgical infections due to rapidly growing Received in revised form 22 July 2017 mycobacteria (RGM). The authors’ experience working with patients with RGM infections who have Accepted 26 July 2017 returned to the USA after traveling abroad for cosmetic surgical procedures is described here. Corresponding Editor: Eskild Petersen, Methods: Patients who developed RGM infections after undergoing cosmetic surgeries abroad and who ?Aarhus, Denmark presented at the Montefiore Medical Center (Bronx, New York, USA) between August 2015 and June 2016 were identified. A review of patient medical records was performed. Keywords: Results: Four patients who presented with culture-proven RGM infections at the sites of recent cosmetic Mycobacterium abscessus complex procedures were identified. -

In Vitro and in Vivo Study of Fosfomycin in Methicillin-Resistant Staphylococcus Aureus Septicaemia
J. Ilyg., Camb. (1980), 96, 419-423 419 Printed in Great Britain In vitro and in vivo study of fosfomycin in methicillin-resistant Staphylococcus aureus septicaemia BY W. Y. LAU, C. H. TEOH-CHAN,* S. T. FAN AND K. F. LAU* Government Surgical Unit, Queen Mary Hospital, and * Department of Microbiology, University of Hong Kong {Received 15 October 19S5; accepted 12 December 19S5) SUMMARY Five hundred strains of methicillin-resistant Staphylococcus aureus were tested against various anti-staphylococcal agents. Vancomycin, fusidic acid and fosfo- mycin were found to be the most effective. Only 1 strain out of 500 was resistant to fosfomycin. Three patients with methicillin-resistant Staphylococcus aureus septicaemia were successfully treated by fosfomycin. We conclude that fosfomycin could be the drug of choice for methicillin-resistant Staphylococcns aureus infection. INTRODUCTION Outbreaks of methicillin-resistant Staphylococcus aureus (MRSA) infection are causing increasing problems in centres all over the world (Thompson & Wenzel, 1982; Editorial, 1983). There has been a gradual increase in the incidence of MRSA in our hospital in Hong Kong from 317 in 1982 (18-7% of all S. aureus isolated) to 511 in 1983 (32-7%) and 833 in 1984 (34-9%). In vitro, resistance of MRSA is often demonstrated to many anti-staphylococcal agents including the penicillins, first-generation cephalosporins, aminoglycosides, erythromycin, clindamycin and chloramphenicol (Peacock et al. 1981). Second- and third-generation cephalosporins also have poor activity in vitro (Thompson, Fisher & Wenzel, 1982) though many MRSA strains have been found to be susceptible to rifampicin, trimethoprim- sulphamethoxazole and fusidic acid (Watanakunakorn, 1983). However, clinical experience of treating serious staphylococeal infection with rifampicin and trimethoprim-siilphamethoxazole is limited (Guenther & Wenzel, 1984) and fully resistant organisms may emerge rapidly after exposure to fusidic acid in vitro and in vivo (Watanakunakorn, 1983). -
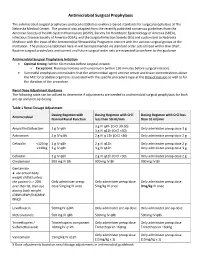
Antimicrobial Surgical Prophylaxis
Antimicrobial Surgical Prophylaxis The antimicrobial surgical prophylaxis protocol establishes evidence-based standards for surgical prophylaxis at The Nebraska Medical Center. The protocol was adapted from the recently published consensus guidelines from the American Society of Health-System Pharmacists (ASHP), Society for Healthcare Epidemiology of America (SHEA), Infectious Disease Society of America (IDSA), and the Surgical Infection Society (SIS) and customized to Nebraska Medicine with the input of the Antimicrobial Stewardship Program in concert with the various surgical groups at the institution. The protocol established here-in will be implemented via standard order sets utilized within One Chart. Routine surgical prophylaxis and current and future surgical order sets are expected to conform to this guidance. Antimicrobial Surgical Prophylaxis Initiation Optimal timing: Within 60 minutes before surgical incision o Exceptions: Fluoroquinolones and vancomycin (within 120 minutes before surgical incision) Successful prophylaxis necessitates that the antimicrobial agent achieve serum and tissue concentrations above the MIC for probable organisms associated with the specific procedure type at the time of incision as well as for the duration of the procedure. Renal Dose Adjustment Guidance The following table can be utilized to determine if adjustments are needed to antimicrobial surgical prophylaxis for both pre-op and post-op dosing. Table 1 Renal Dosage Adjustment Dosing Regimen with Dosing Regimen with CrCl Dosing Regimen with -

Antimicrobial Stewardship Guidance
Antimicrobial Stewardship Guidance Federal Bureau of Prisons Clinical Practice Guidelines March 2013 Clinical guidelines are made available to the public for informational purposes only. The Federal Bureau of Prisons (BOP) does not warrant these guidelines for any other purpose, and assumes no responsibility for any injury or damage resulting from the reliance thereof. Proper medical practice necessitates that all cases are evaluated on an individual basis and that treatment decisions are patient-specific. Consult the BOP Clinical Practice Guidelines Web page to determine the date of the most recent update to this document: http://www.bop.gov/news/medresources.jsp Federal Bureau of Prisons Antimicrobial Stewardship Guidance Clinical Practice Guidelines March 2013 Table of Contents 1. Purpose ............................................................................................................................................. 3 2. Introduction ...................................................................................................................................... 3 3. Antimicrobial Stewardship in the BOP............................................................................................ 4 4. General Guidance for Diagnosis and Identifying Infection ............................................................. 5 Diagnosis of Specific Infections ........................................................................................................ 6 Upper Respiratory Infections (not otherwise specified) .............................................................................. -

Susceptibility of Pseudomonas Cepacia Isolated from Children with Cystic Fibrosis1
003 1-3998/86/2011-1174$02.00/0 PEDIATRIC RESEARCH Vol. 20, No. 1 1, 1986 Copyright O 1986 International Pediatric Research Foundation, Inc. Printed in (I.S.A. Decreased Baseline P-Lactamase Production and Inducibility associated with Increased Piperacillin Susceptibility of Pseudomonas cepacia Isolated from Children with Cystic Fibrosis1 CLAUDIO CHIESA,~PAULINE H. LABROZZI, AND STEPHEN C. ARONOFF Department of Pediatrics [C.C., P.H.L., S.C.A.], Case- Western Reserve University School ofMedicine and the Division of Pediatric Infectious Diseases [S.C.A.], Rainbow Babies and Children's Hospital, Cleveland, Ohio ABSTRACT. The incidence of pulmonary infections in creased since 1978 and is now recovered from 20% of patients children with cystic fibrosis caused by Pseudomonas ce- in some centers (3, 4). pacia, an organism which may possess an inducible 8- Sputum isolates of P. cepacia from children with cystic fibrosis lactamase, has increased since 1978. Seven of 13 sputum are resistant to most P-lactam agents in vitro. In an in vitro study isolates of P. cepacia from children with cystic fibrosis comparing the susceptibilities of 62 consecutive sputum isolates, were classified as inducible by quantitative enzyme produc- concentrations of 64 pg/ml of aztreonam and piperacillin were tion following preincubation with 100, 200, or 400 pg/ml required to inhibit 79 and 87.1 % of the bacterial population, of cefoxitin. The recovery of inducible strains tended to be respectively; 90% of the isolates were inhibited by 8 pg/ml of associated with recent ceftazidime therapy. Susceptibility ceftazidime (5). In vitro, ceftazidime has proven more active to aztreonam, ceftazidime, and piperacillin alone or com- against P. -

Thienamycin (Imipenem)
1520 THE JOURNAL OF ANTIBIOTICS OCT. 1989 CONVERSION OF OA-6129 B2 INTO OA-6129 B2, a fermentation carbapenem product, (5i?,6^)-6-[(i?)- l -FLUOROETHYL]-7- into optically active 88617, an 8-fluorinated OXO- 3 -[(7V,Ar,Ar-TRIM ETHYL- carbapenem derivative which improved antimicro- CARBAMIMIDOYL)METHYL]THIO- bial activity and dehydropeptidase-I stability. l -AZABICYCLO[3.2.0]HEPT-2-ENE- Treatment of OA-6129B2 sodium salt 1 with 2-CARBOXYLIC ACID (88617)t /7-nitrobenzyl (PNB) bromide or pivaloyloxymethyl chloride in DMFgave /?NB ester 2 or pivaloyl- Takeo Yoshioka, AzumaWatanabe, oxymethyl (POM) ester 3 of OA-6129 B2 (Fig. 1). Noritaka Chida and Yasuo Fukagawa The two hydroxyl groups in the pantoyl moiety of OA-6129B2 were protected by isopropylidena- Central Research Laboratories, tion for differentiation from the C-8 hydroxyl Sanraku Incorporated, 4-9-1 Johnan, Fujisawa 251, Japan group. The protected compound4 or 5 was allowed to (Received for publication May 25, 1989) react with 1.2 equiv of diethylaminosulfur trifluoride (DAST)11} at -68°C in methylene chloride for 10 minutes. After the reaction mixture was diluted with Since the epoch-making discovery of thienamy- methylene chloride and washed with a saturated cin2), a numberof extensive studies have been carried aqueous sodium bicarbonate solution, silica gel out on the development of clinically useful car- column chromatography of the organic layer pro- bapenem compounds. As a result, formimidoyl- vided the desired fluorinated compound 6 or 7 thienamycin (imipenem) has recently been put on having a fluorine atom in the R configuration at C-8 the market as a combination drug (Primaxin)3) in 43 or 49% yield, respectively. -

The Serpintine Solution
& Experim l e ca n i t in a l l C C f a Journal of Clinical & Experimental o r d l i a o n l o r g u Lucas et al., J Clin Exp Cardiolog 2017, 8:1 y o J Cardiology ISSN: 2155-9880 DOI: 10.4172/2155-9880.1000e150 Editorial Open Access The Serpintine Solution Alexandra Lucas, MD, FRCP(C)1,2,*, Sriram Ambadapadi, PhD1, Brian Mahon, PhD3, Kasinath Viswanathan, PhD4, Hao Chen, MD, PhD5, Liying Liu, MD6, Erbin Dai, MD6, Ganesh Munuswami-Ramanujam, PhD7, Jacek M. Kwiecien, DVM, MSc, PhD8, Jordan R Yaron, PhD1, Purushottam Shivaji Narute, BVSc & AH, MVSc, PhD1,9, Robert McKenna, PhD10, Shahar Keinan, PhD11, Westley Reeves, MD, PhD12, Mark Brantly, MD, PhD13, Carl Pepine, MD, FACC14 and Grant McFadden, PhD1 1Center for Personalized Diagnostics, Biodesign Institute, Arizona State University, Tempe AZ, USA 2Saint Josephs Hospital, Dignity Health Phoenix, Phoenix, AZ, USA 3NIH/ NIDDK, Bethesda MD, USA 4Zydus Research Centre, Ahmedabad, India 5Department of tumor surgery, The Second Hospital of Lanzhou University, Lanzhou, Gansu, P.R.China 6Beth Israel Deaconess Medical Center, Harvard, Boston, MA, USA 7Interdisciplinary Institute of Indian system of Medicine (IIISM), SRM University, Chennai, Tamil Nadu, India 8MacMaster University, Hamilton, ON, Canada 9Critical Care Medicine Department, Clinical Center, National Institutes of Health, Bethesda MD, USA 10Department of Molecular Genetics and Microbiology, University of Florida, Gainesville, FL, USA 11Cloud Pharmaceuticals, Durham, North Carolina, USA 12Division of Rheumatology, University of Florida, -

Australian Public Assessment Refport for Ceftaroline Fosamil (Zinforo)
Australian Public Assessment Report for ceftaroline fosamil Proprietary Product Name: Zinforo Sponsor: AstraZeneca Pty Ltd May 2013 Therapeutic Goods Administration About the Therapeutic Goods Administration (TGA) • The Therapeutic Goods Administration (TGA) is part of the Australian Government Department of Health and Ageing, and is responsible for regulating medicines and medical devices. • The TGA administers the Therapeutic Goods Act 1989 (the Act), applying a risk management approach designed to ensure therapeutic goods supplied in Australia meet acceptable standards of quality, safety and efficacy (performance), when necessary. • The work of the TGA is based on applying scientific and clinical expertise to decision- making, to ensure that the benefits to consumers outweigh any risks associated with the use of medicines and medical devices. • The TGA relies on the public, healthcare professionals and industry to report problems with medicines or medical devices. TGA investigates reports received by it to determine any necessary regulatory action. • To report a problem with a medicine or medical device, please see the information on the TGA website <http://www.tga.gov.au>. About AusPARs • An Australian Public Assessment Record (AusPAR) provides information about the evaluation of a prescription medicine and the considerations that led the TGA to approve or not approve a prescription medicine submission. • AusPARs are prepared and published by the TGA. • An AusPAR is prepared for submissions that relate to new chemical entities, generic medicines, major variations, and extensions of indications. • An AusPAR is a static document, in that it will provide information that relates to a submission at a particular point in time. • A new AusPAR will be developed to reflect changes to indications and/or major variations to a prescription medicine subject to evaluation by the TGA. -

Comparative Ceftaroline Activity Tested Against Staphylococcus Aureus Associated with Acute Bacterial Skin and Skin Structure Infection from a Tertiary Care Center
RESEARCH PAPER Medical Science Volume : 5 | Issue : 4 | April 2015 | ISSN - 2249-555X Comparative Ceftaroline Activity Tested Against Staphylococcus Aureus Associated with Acute Bacterial Skin and Skin Structure Infection from A Tertiary Care Center KEYWORDS MRSA, Ceftaroline, vancomycin Dr. Umamageswari S. S. M. Dr. S. Habeeb Mohammed Dr. Shameem Banu Associate Professor- Microbiology Associate Professor-Surgery Professor and HOD- Microbiology Chettinad Hospital & Research Chettinad Hospital & Research Chettinad Hospital & Research Insitute Insitute Insitute Dr. Jayanthi S. Associate Professor- Microbiology Chettinad Hospital & Research Insitute ABSTRACT Methicillin resistant Staphylococcus aureus (MRSA), commonest cause of acute bacterial skin and skin structure infection (ABSSSI) has only limited treatment options like vancomycin, Linezolid and teicoplanin. Staphylococcus aureus with reduced susceptibilities to vancomycin is developing nowadays among nosocomial infec- tion. A newest cephalosporin – Ceftaroline has received FDA approval for the treatment of ABSSSI recently has unique activity against MRSA. The aim of the study is to know the prevalence of Staphylococcus aureus in ABSSSI patients and to evaluate the in vitro activity of Ceftaroline, in comparison with other drugs. Minimum Inhibitory Concentration for vancomycin and Ceftaroline were determined by E-test strips. Out of 235 Staphylococcus aureus isolated from ABSSSI patient 32% were MRSA. No vancomycin resistant strain isolated in our study. In vitro, Ceftaroline control the growth of MRSA very effectively at 2mg/ml itself. Ceftaroline is a most welcomed drug in the treatment of MRSA. INTRODUCTION: MATERIALS AND METHODS: Complicated skin and skin structure infection (CSSSIs), now This study was carried out in Chettinad health city and known by the new US FDA as Acute bacterial skin and skin Research Institute, a tertiary care center hospital (Chen- structure infections (ABSSSI) are the most common infec- nai) between April 2013 and May 2014. -

Penicillin Allergy Guidance Document
Penicillin Allergy Guidance Document Key Points Background Careful evaluation of antibiotic allergy and prior tolerance history is essential to providing optimal treatment The true incidence of penicillin hypersensitivity amongst patients in the United States is less than 1% Alterations in antibiotic prescribing due to reported penicillin allergy has been shown to result in higher costs, increased risk of antibiotic resistance, and worse patient outcomes Cross-reactivity between truly penicillin allergic patients and later generation cephalosporins and/or carbapenems is rare Evaluation of Penicillin Allergy Obtain a detailed history of allergic reaction Classify the type and severity of the reaction paying particular attention to any IgE-mediated reactions (e.g., anaphylaxis, hives, angioedema, etc.) (Table 1) Evaluate prior tolerance of beta-lactam antibiotics utilizing patient interview or the electronic medical record Recommendations for Challenging Penicillin Allergic Patients See Figure 1 Follow-Up Document tolerance or intolerance in the patient’s allergy history Consider referring to allergy clinic for skin testing Created July 2017 by Macey Wolfe, PharmD; John Schoen, PharmD, BCPS; Scott Bergman, PharmD, BCPS; Sara May, MD; and Trevor Van Schooneveld, MD, FACP Disclaimer: This resource is intended for non-commercial educational and quality improvement purposes. Outside entities may utilize for these purposes, but must acknowledge the source. The guidance is intended to assist practitioners in managing a clinical situation but is not mandatory. The interprofessional group of authors have made considerable efforts to ensure the information upon which they are based is accurate and up to date. Any treatments have some inherent risk. Recommendations are meant to improve quality of patient care yet should not replace clinical judgment. -

Understanding Drug-Drug Interactions Due to Mechanism-Based Inhibition in Clinical Practice
pharmaceutics Review Mechanisms of CYP450 Inhibition: Understanding Drug-Drug Interactions Due to Mechanism-Based Inhibition in Clinical Practice Malavika Deodhar 1, Sweilem B Al Rihani 1 , Meghan J. Arwood 1, Lucy Darakjian 1, Pamela Dow 1 , Jacques Turgeon 1,2 and Veronique Michaud 1,2,* 1 Tabula Rasa HealthCare Precision Pharmacotherapy Research and Development Institute, Orlando, FL 32827, USA; [email protected] (M.D.); [email protected] (S.B.A.R.); [email protected] (M.J.A.); [email protected] (L.D.); [email protected] (P.D.); [email protected] (J.T.) 2 Faculty of Pharmacy, Université de Montréal, Montreal, QC H3C 3J7, Canada * Correspondence: [email protected]; Tel.: +1-856-938-8697 Received: 5 August 2020; Accepted: 31 August 2020; Published: 4 September 2020 Abstract: In an ageing society, polypharmacy has become a major public health and economic issue. Overuse of medications, especially in patients with chronic diseases, carries major health risks. One common consequence of polypharmacy is the increased emergence of adverse drug events, mainly from drug–drug interactions. The majority of currently available drugs are metabolized by CYP450 enzymes. Interactions due to shared CYP450-mediated metabolic pathways for two or more drugs are frequent, especially through reversible or irreversible CYP450 inhibition. The magnitude of these interactions depends on several factors, including varying affinity and concentration of substrates, time delay between the administration of the drugs, and mechanisms of CYP450 inhibition. Various types of CYP450 inhibition (competitive, non-competitive, mechanism-based) have been observed clinically, and interactions of these types require a distinct clinical management strategy. This review focuses on mechanism-based inhibition, which occurs when a substrate forms a reactive intermediate, creating a stable enzyme–intermediate complex that irreversibly reduces enzyme activity. -

Profile Profile Profile Uses and Administration
Gramicidin/lmipenem 309 Graneodin N; Graneodin; Gripaben Caramelos; Kenacomb; t Incompatibility and stability. Intipenem is unstable at Nasomicina; Neo Coltirot; Pantometilt; Proetztotal; Tosedrin �-�P.�.�� ����---········································································· alkaline or acidic pH and the commercially available injec (details are given in Volume B) NF; Austral.: Kenacomb; Neosporint; Otocomb Otic; Otodex; ProprielaryPreparations tion of intipenem with cilastatin sodium for intravenous Sofradex; Soframycint; Austria: Volon A antibiotikahaltigt; V alpeda. use is buffered to provide, when reconstituted, a solution Belg. : Mycolog; Polyspectran Granticidinet; Braz.: Fonergin; Single-ingredient Preparations. UK: with pH 6.5 to 7.5. Licensed product information advises Londerm-Nt; Mud; Neolon D; Omcilon-A M; Oncibel; Oncileg; against mixing with other antibacterials. Onciplust; Canad.: Ak Spor; Antibiotic Cream; Antibiotic Plus; Complete Antibiotic Ointment; Diosporint; Neosporin; Opti lbafloxacin (BAN, USAN, r/NN) References. cort; Optimyxin Plus; Optimyxin; Polysporin Complete; Poly 1. Bigley FP, et al. Compatibility of imipenem-dlastatin sodium with Am Hosp Phann 1986; 43: 2803- sporin For Kids; Polysporin Plus Pain Relief; Polysporin Triple commonly used intravenous solutions. 9. J Antibiotic; Polysporin; Polytopic; ratio-Tria comb; Sofracort; 2. Smith GB, et al. Stability and kinetics of degradation of imipenem in Soframycin; Soframycin; Triple Antibiotic Ointment; Viaderm Pharm 1990; 79: 732-40. aqueous