The Occurrence of Red and Yellow Autumn Leaves Explained by Regional Differences in Insolation and Temperature
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Cally Plant List a ACIPHYLLA Horrida
Cally Plant List A ACIPHYLLA horrida ACONITUM albo-violaceum albiflorum ABELIOPHYLLUM distichum ACONITUM cultivar ABUTILON vitifolium ‘Album’ ACONITUM pubiceps ‘Blue Form’ ACAENA magellanica ACONITUM pubiceps ‘White Form’ ACAENA species ACONITUM ‘Spark’s Variety’ ACAENA microphylla ‘Kupferteppich’ ACONITUM cammarum ‘Bicolor’ ACANTHUS mollis Latifolius ACONITUM cammarum ‘Franz Marc’ ACANTHUS spinosus Spinosissimus ACONITUM lycoctonum vulparia ACANTHUS ‘Summer Beauty’ ACONITUM variegatum ACANTHUS dioscoridis perringii ACONITUM alboviolaceum ACANTHUS dioscoridis ACONITUM lycoctonum neapolitanum ACANTHUS spinosus ACONITUM paniculatum ACANTHUS hungaricus ACONITUM species ex. China (Ron 291) ACANTHUS mollis ‘Long Spike’ ACONITUM japonicum ACANTHUS mollis free-flowering ACONITUM species Ex. Japan ACANTHUS mollis ‘Turkish Form’ ACONITUM episcopale ACANTHUS mollis ‘Hollard’s Gold’ ACONITUM ex. Russia ACANTHUS syriacus ACONITUM carmichaelii ‘Spätlese’ ACER japonicum ‘Aconitifolium’ ACONITUM yezoense ACER palmatum ‘Filigree’ ACONITUM carmichaelii ‘Barker’s Variety’ ACHILLEA grandifolia ACONITUM ‘Newry Blue’ ACHILLEA ptarmica ‘Perry’s White’ ACONITUM napellus ‘Bergfürst’ ACHILLEA clypeolata ACONITUM unciniatum ACIPHYLLA monroi ACONITUM napellus ‘Blue Valley’ ACIPHYLLA squarrosa ACONITUM lycoctonum ‘Russian Yellow’ ACIPHYLLA subflabellata ACONITUM japonicum subcuneatum ACONITUM meta-japonicum ADENOPHORA aurita ACONITUM napellus ‘Carneum’ ADIANTUM aleuticum ‘Japonicum’ ACONITUM arcuatum B&SWJ 774 ADIANTUM aleuticum ‘Miss Sharples’ ACORUS calamus ‘Argenteostriatus’ -
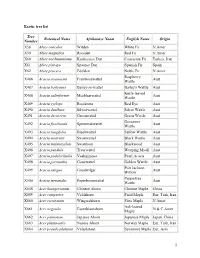
1 Exotic Tree List Tree Number Botanical Name Afrikaanse Naam
Exotic tree list Tree Botanical Name Afrikaanse Naam English Name Origin Number X58 Abies concolor Witden White Fir N.Amer X59 Abies magnifica Rooiden Red Fir N.Amer X60 Abies nordmanniana Kaukasiese Den Caucasian Fir Turkey, Iran X61 Abies pinsapo Spaanse Den Spanish Fir Spain X62 Abies procera Edelden Noble Fir N.Amer Raspberry X486 Acacia acuminata Frambosewattel Aust Wattle X487 Acacia baileyana Bailey-se-wattel Bailey's Wattle Aust Knife-leaved X488 Acacia cultriformis Mesblaarwattel Aust Wattle X489 Acacia cyclops Rooikrans Red Eye Aust X490 Acacia dealbata Silwerwattel Silver Wattle Aust X491 Acacia decurrens Groenwattel Green Wattle Aust Gossamer X492 Acacia floribunda Spinnerakwattel Aust Wattle X493 Acacia longifolia Bleekwattel Sallow Wattle Aust X494 Acacia mearnsii Swartwattel Black Wattle Aust X495 Acacia melanoxylon Swarthout Blackwood Aust X496 Acacia pendula Treurwattel Weeping Myall Aust X497 Acacia podalyriifolia Vaalmimosa Pearl Acacia Aust X498 Acacia pycnantha Gouewattel Golden Wattle Aust Port Jackson X499 Acacia saligna Goudwilger Aust Willow Peppertree X500 Acacia terminalis Peperboomwattel Aust Wattle X658 Acer buergerianum Chinese Ahorn Chinese Maple China X659 Acer campestre Veldahorn Field Maple Eur, Turk, Iran X660 Acer circinatum Wingerdahorn Vine Maple N Amer Ash-leaved X661 Acer negundo Essenblaarahorn N & C Amer Maple X662 Acer palmatum Japanse Ahorn Japanese Maple Japan, China X663 Acer platanoides Noorse Ahorn Norway Maple Eur, Turk, Iran X664 Acer pseudo-platanus Valsplataan Sycamore Maple Eur, Asia -

Ecological Basis for the Control of Gunnera Tinctoria in Sao Miguel
Second International Weed Control Congress Copenhagen 1996 . Ecological basis for the control of Gunnera tinctoria in Sao Miguel . Island By L sn.. v A, JT A V ARES and A PENA Departamento de Biologia, Universidade dos A(:ores, PT-9500 Ponta Dell?lUia, Portugal, E-mail [email protected] Summary Gunnera tinctoria, an herbaceous plant from South America, is naturalised in Sao Miguel island (Azores) . .In this research an ecologically based strategy for G. tinctoria control is suggested. Infestation structure, altitudinal range, associated plants, phenology and natural enemies were studied. G. tillctoria was found from \00 to 900 m of altitude, in plane or highly sloped terrain, on rich soil or gravel, in roadsides, trails, and water streams. Infestation foci were found at 40 Krn from introduction site. Populations consisted of isolated or small groups of plants, with reduced cover, associated with other weeds. According to three plant invaders classification systems this plant presents several negative characters in terms of conservation: high seed production, vegetative reproduction, high impact on the landscape. invasion of natural vegetation. Priority of control should be given to satellite populations in high conservation ·value sites. Control of heavier infestations will need a persistent and global approach. Introduction Natural populations of Gunnera sp. are restricted to super-humid areas with heavy rainfall; they prefer high altitudes and open or lightly shaded areas, and are often pioneers on bare land (Bergman et al., 1992). The Gunneracea includes the genus Gunnera L., with terrestrial, rhizomatous, perennial herbs, sometimes gigantic, from tropical and warm temperate regions. The larger kinds are grown for the striking effect of their enormous leaves, the smaller forms. -

Laurisilva of Madeira Portugal
LAURISILVA OF MADEIRA PORTUGAL The Laurisilva of Madeira is the largest surviving relict of a virtually extinct laurel forest type once widespread in Europe. It is still 90% primary forest and is a centre of plant diversity, containing a unique suite of rare and relict plants and animals, especially endemic bryophytes, ferns, vascular plants, animals such as the Madeiran long-toed pigeon and a very rich invertebrate fauna. COUNTRY Portugal NAME Laurisilva of Madeira NATURAL WORLD HERITAGE SITE 1999: Inscribed on the World Heritage List under Natural Criteria ix and x. STATEMENT OF OUTSTANDING UNIVERSAL VALUE The UNESCO World Heritage Committee adopted the following Statement of Outstanding Universal Value at the time of inscription: Brief Synthesis The Laurisilva of Madeira, within the Parque Natural da Madeira (Madeira Natural Park) conserves the largest surviving area of primary laurel forest or "laurisilva", a vegetation type that is now confined to the Azores, Madeira and the Canary Islands. These forests display a wealth of ecological niches, intact ecosystem processes, and play a predominant role in maintaining the hydrological balance on the Island of Madeira. The property has great importance for biodiversity conservation with at least 76 vascular plant species endemic to Madeira occurring in the property, together with a high number of endemic invertebrates and two endemic birds including the emblematic Madeiran Laurel Pigeon. Criterion (ix): The Laurisilva of Madeira is an outstanding relict of a previously widespread laurel forest type, which covered much of Southern Europe 15-40 million years ago. The forest of the property completely covers a series of very steep, V-shaped valleys leading from the plateau and east-west ridge in the centre of the island to the north coast. -

“Pico Da Dara” Natural Reserve (S. Miguel Island, Azores Islands, Portugal) Using Ikonos Satellite Imagery
XIII Congreso de la Sociedad Española de Malherbología, La Laguna 2011 VEGETATION MAPPING OF “PICO DA DARA” NATURAL RESERVE (S. MIGUEL ISLAND, AZORES ISLANDS, PORTUGAL) USING IKONOS SATELLITE IMAGERY A. Gil CITA-A (Azorean Biodiversity Group), Department of Biology, University of the Azores, 9501- 801 Ponta Delgada, Portugal. [email protected] Resumen: La flora vascular de la isla de S.Miguel (Archipiélago de las Azores, Portugal) es constituída por cerca de 1000 taxa y es largamente dominada por especies exoticas. La rapida propagación de especies exoticas invasoras muy agresivas como Pittosporum undulatum Vent. y Clethra arborea Aiton está causando graves problemas de conservación en ese ecosistema insular. Este artigo tiene como objectivo evaluar la efectividad de imágenes satelitales de alta resolución espacial IKONOS para cartografiar la vegetación de la Reserva Natural de Pico da Vara, usando 4 diferentes técnicas supervisionadas de clasificación (paramétricas y no paramétricas). Los resultados obtenidos han demostrado que el uso de imágenes IKONOS puede constituir un método efectivo para la caracterización, monitoreo y evaluación de los ecosistemas terrestres en las Azores. Palabras Clave: teledetección, Cartografía de Vegetación, Alta Resolución. INTRODUCTION S. Miguel Island's vascular plant flora (Archipelago of the Azores, Portugal) is largely dominated by non-indigenous taxa. However, existing indigenous vascular plant taxa are particularly important because they compose a very valuable ecosystem, the Azorean Laurel Forest. One of its most significant areas is the core of Pico da Vara / Ribeira do Guilherme Special Protected Area, in the former Natural Reserve of Pico da Vara (the case-study area), located in the mountain complex of Serra da Tronqueira. -

Linking Dendrometry and Dendrochronology in the Dominant Azorean Tree Laurus Azorica (Seub.) Franco
Article Linking Dendrometry and Dendrochronology in the Dominant Azorean Tree Laurus azorica (Seub.) Franco Bárbara Matos 1,2, Lurdes Borges Silva 1,2,*, Ricardo Camarinho 1,3, Armindo S. Rodrigues 1,3, Ruben Rego 1,2 , Mariana Câmara 1 and Luís Silva 1,2 1 Faculdade de Ciências e Tecnologia, Universidade dos Açores, Rua Mãe de Deus 13A, 9501–855 Ponta Delgada, Portugal 2 InBIO, Rede de Investigação em Biodiversidade e Biologia Evolutiva, Laboratório Associado, CIBIO-Açores, Universidade dos Açores, Apartado 1422, 9501-801 Ponta Delgada, Portugal 3 IVAR, Institute of Volcanology and Risks Assessment, University of the Azores, 9501-801 Ponta Delgada, Portugal * Correspondence: [email protected]; Tel.: +351-296-650-105 Received: 14 May 2019; Accepted: 21 June 2019; Published: 27 June 2019 Abstract: As in many archipelagos, the Azorean primary forest was largely cleared and replaced by secondary forest and grassland, the Azorean tree Laurus azorica (Seub.) Franco being one of the dominant trees in the remaining natural forests. Dendrochronological and dendrometric studies in the Azores mainly focused on non-indigenous trees, either used for timber (e.g., Cryptomeria japonica D. Don) or considered as invasive (Pittosporum undulatum Vent.). Therefore, this study aims to describe the growth ring anatomy of L. azorica, and to understand the relationship between dendrometric traits (e.g., trunk diameter at breast height; tree height), and the number of growth rings. Growth ring anatomy was accessed by wood anatomical preparation of microcore samples while tree age estimation was based on growth ring counts in wood cores taken at breast height and at base. -

Patterns of Colonization and Species Distribution for Azorean Arthropods: Evolution, Diversity, Rarity and Extinction
AÇOREANA, 2011, Suplemento 7: 93-123 PATTERNS OF COLONIZATION AND SPECIES DISTRIBUTION FOR AZOREAN ARTHROPODS: EVOLUTION, DIVERSITY, RARITY AND EXTINCTION Paulo A.V. Borges1, Clara Gaspar1, Ana M.C. Santos1, Sérvio Pontes Ribeiro1,2, Pedro Cardoso1,3, Kostas A. Triantis1,4 & Isabel R. Amorim1,5 1 Azorean Biodiversity Group (CITA-A), Departamento de Ciências Agrárias, Universidade dos Açores, Pico da Urze, 9700-042 Angra do Heroísmo, Portugal e-mail: [email protected] 2 Universidade Federal de Ouro Preto, Instituto de Ciências Exatas e Biológicas, Lab. Ecologia Evolutiva de Herbívoros de Dossel/DEBIO, campus Morro do Cruzeiro, 35400-000, Ouro Preto, MG, Brasil 3Smithsonian Institution, National Museum of Natural History, Washington, DC, USA Department of Biology, Texas State University, San Marcos, TX, USA 4Biodiversity Research Group, Oxford University Centre for the Environment, South Parks Road, Oxford, OX1 3QY, UK 5School of Biological Sciences, University of East Anglia, Norwich NR4 7TJ, England, UK ABSTRACT Here we address a list of questions based on long-term ecological and biogeographical studies performed in the Azores, a remote volcanic oceanic archipelago composed by nine islands. The target group are the arthropods, and the main habitat the Laurisilva, the Azorean native forest. Diversification of Azorean arthropod species is affected by island age, area and isolation. However, results obtained for over a decade show that distinct groups are differently affected by these factors, which has lead to the extreme diverse distribution patterns currently observed. Spatial distribution of arthropods in each island may be interpreted as caused by a typical “mass effect”, with many species following a “source-sink” dynamics. -

Gardens and Stewardship
GARDENS AND STEWARDSHIP Thaddeus Zagorski (Bachelor of Theology; Diploma of Education; Certificate 111 in Amenity Horticulture; Graduate Diploma in Environmental Studies with Honours) Submitted in fulfilment of the requirements for the degree of Doctor of Philosophy October 2007 School of Geography and Environmental Studies University of Tasmania STATEMENT OF AUTHENTICITY This thesis contains no material which has been accepted for any other degree or graduate diploma by the University of Tasmania or in any other tertiary institution and, to the best of my knowledge and belief, this thesis contains no copy or paraphrase of material previously published or written by other persons, except where due acknowledgement is made in the text of the thesis or in footnotes. Thaddeus Zagorski University of Tasmania Date: This thesis may be made available for loan or limited copying in accordance with the Australian Copyright Act of 1968. Thaddeus Zagorski University of Tasmania Date: ACKNOWLEDGEMENTS This thesis is not merely the achievement of a personal goal, but a culmination of a journey that started many, many years ago. As culmination it is also an impetus to continue to that journey. In achieving this personal goal many people, supervisors, friends, family and University colleagues have been instrumental in contributing to the final product. The initial motivation and inspiration for me to start this study was given by Professor Jamie Kirkpatrick, Dr. Elaine Stratford, and my friend Alison Howman. For that challenge I thank you. I am deeply indebted to my three supervisors Professor Jamie Kirkpatrick, Dr. Elaine Stratford and Dr. Aidan Davison. Each in their individual, concerted and special way guided me to this omega point. -

Peloptulus Borgesi
The IUCN Red List of Threatened Species™ ISSN 2307-8235 (online) IUCN 2020: T119441267A119441683 Scope(s): Global Language: English Peloptulus borgesi Assessment by: Nunes, R. & Borges, P.A.V. View on www.iucnredlist.org Citation: Nunes, R. & Borges, P.A.V. 2020. Peloptulus borgesi. The IUCN Red List of Threatened Species 2020: e.T119441267A119441683. https://dx.doi.org/10.2305/IUCN.UK.2020- 3.RLTS.T119441267A119441683.en Copyright: © 2020 International Union for Conservation of Nature and Natural Resources Reproduction of this publication for educational or other non-commercial purposes is authorized without prior written permission from the copyright holder provided the source is fully acknowledged. Reproduction of this publication for resale, reposting or other commercial purposes is prohibited without prior written permission from the copyright holder. For further details see Terms of Use. The IUCN Red List of Threatened Species™ is produced and managed by the IUCN Global Species Programme, the IUCN Species Survival Commission (SSC) and The IUCN Red List Partnership. The IUCN Red List Partners are: Arizona State University; BirdLife International; Botanic Gardens Conservation International; Conservation International; NatureServe; Royal Botanic Gardens, Kew; Sapienza University of Rome; Texas A&M University; and Zoological Society of London. If you see any errors or have any questions or suggestions on what is shown in this document, please provide us with feedback so that we can correct or extend the information provided. THE IUCN RED LIST OF THREATENED SPECIES™ Taxonomy Kingdom Phylum Class Order Family Animalia Arthropoda Arachnida Oribatida Phenopelopidae Scientific Name: Peloptulus borgesi Pérez-Íñigo & Pérez-Íñigo Jr., 1996 Assessment Information Red List Category & Criteria: Data Deficient ver 3.1 Year Published: 2020 Date Assessed: March 31, 2018 Justification: Peloptulus borgesi is an endemic species of the Azores (Portugal), described from the island of S. -

Status Assessment of the Critically Endangered Azores Bullfinch Pyrrhula Murina
Bird Conservation International, page 1 of 13. © BirdLife International, 2011 doi:10.1017/S0959270911000025 Status assessment of the Critically Endangered Azores Bullfinch Pyrrhula murina RICARDO S. CEIA, JAIME A. RAMOS, RUBEN H. HELENO, GEOFF M. HILTON and TIAGO A. MARQUES Summary The Azores Bullfinch is endemic to the island of São Miguel (Azores, Portugal). Its status was uplisted to Critically Endangered in 2005 on the basis of an extremely small and declining 2 population that was considered to be restricted to a very small mountain range (43 km ), in a single location, within which the spread of invasive plants constituted a threat to habitat quality. Nevertheless, information was mostly inferred, or the product of, non-systematic studies. In order to carry out a complete assessment of the conservation status we analysed: (i) population trend, calculated from annual monitoring 1991–2008, (ii) population size, and (iii) range size, obtaining estimates in a single morning study in 2008 involving the simultaneous participation of 48 observers. Contrary to previous inferences, the population is no longer decreasing, although quality of laurel forest habitat continues to decline due to the persistent threat of invasive species. Population size (mean 6 SE) was estimated at 1,064 6 304 individuals using distance sampling methods, although the estimate was very sensitive to the survey method used. Range size 2 2 estimates (extent of occurrence and area of occupancy) were 144 km and 83 km respectively. Given the present information, we propose the downlisting of Azores Bullfinch to Endangered on the IUCN Red List. Introduction The Azores Bullfinch Pyrrhula murina is one of the only two ‘Critically Endangered’ bird species that breed in Europe (BirdLife International 2009). -

Roridula Gorgonias and a Non-Carnivorous Relative, Clethra Arborea
Mathematisch-Naturwissenschaftliche Fakultät Stefanie Hartmann | Michaela Preick | Silke Abelt | André Scheffel | Michael Hofreiter Annotated genome sequences of the carnivorous plant Roridula gorgonias and a non-carnivorous relative, Clethra arborea Suggested citation referring to the original publication: BMC Research Notes 13 (2020), Art. 426 DOI https://doi.org/10.1186/s13104-020-05254-4 ISSN 1756-0500 Postprint archived at the Institutional Repository of the Potsdam University in: Postprints der Universität Potsdam : Mathematisch-Naturwissenschaftliche Reihe 1141 ISSN: 1866-8372 https://nbn-resolving.org/urn:nbn:de:kobv:517-opus4-503752 DOI: https://doi.org/10.25932/publishup-50375 Hartmann et al. BMC Res Notes (2020) 13:426 https://doi.org/10.1186/s13104-020-05254-4 BMC Research Notes RESEARCH NOTE Open Access Annotated genome sequences of the carnivorous plant Roridula gorgonias and a non-carnivorous relative, Clethra arborea Stefanie Hartmann1* , Michaela Preick1, Silke Abelt1, André Schefel2 and Michael Hofreiter1 Abstract Objective: Plant carnivory is distributed across the tree of life and has evolved at least six times independently, but sequenced and annotated nuclear genomes of carnivorous plants are currently lacking. We have sequenced and structurally annotated the nuclear genome of the carnivorous Roridula gorgonias and that of a non-carnivorous rela- tive, Madeira’s lily-of-the-valley-tree, Clethra arborea, both within the Ericales. This data adds an important resource to study the evolutionary genetics of plant carnivory across angiosperm lineages and also for functional and systematic aspects of plants within the Ericales. Results: Our assemblies have total lengths of 284 Mbp (R. gorgonias) and 511 Mbp (C. arborea) and show high BUSCO scores of 84.2% and 89.5%, respectively. -

Eapr. 28, Apl~In 28, 1858. James Powrie, Esq., Reswallie, Near Forfar
Downloaded from http://jgslegacy.lyellcollection.org/ at University •~0 PROCEEDII~IGSCollege OF London THE GEOLOGICAL on July 2, 2016 SOCIETY. EApr. 28, APl~In 28, 1858. James Powrie, Esq., Reswallie, near Forfar, Marcus ttuish, Esq., Castle Donnington, Derby, l~enry D. M. Spence, Esq., Hyde Park Square, and Parkin Jeffcock, Esq., Derby, were elected Fellows. The following communications were read :-- 1. On some ¥EGETABIm RP.~AI~S from MADEIRA. By C~rA~nF.S J. F. Bvs~cav, Esq., F.R.S., F.G.S. TEE vegetable remains procured by Sir Charles Lyell and M. Hartung from the leaf-bed which they discovered* in the ravine of S. Jorge, in the Island of Madeira, were entrnsted to me for examination ; and I propose now to lay before the Geological Society the observations I have been able to make upon them. I have examined 140 speci- mens, by far the greatest part of them in a very imperfect state, mostly small fragments, often quite lmdeterminable, and, even when most perfect, no more than single detached leaves, of which, how- ever, the veins and margin are often very well displayed. Dico- tyledonous leaves predominate, but arc intermixed with numerous remains of Ferns, always, however, in small fragments, insufficient to give any idea of the general form of the frond. The very fragmentary and incomplete condition of the remains of Ferns in this bed is indeed striking. Something of it is attributable to the natnre of the stone, which is by no means fissile, but breaks quite irregularly; but something also would seem to be owing to the original conditions of the deposit.